Pascal Ribéreau-Gayon - Handbook of Enology - Volume 1
Здесь есть возможность читать онлайн «Pascal Ribéreau-Gayon - Handbook of Enology - Volume 1» — ознакомительный отрывок электронной книги совершенно бесплатно, а после прочтения отрывка купить полную версию. В некоторых случаях можно слушать аудио, скачать через торрент в формате fb2 и присутствует краткое содержание. Жанр: unrecognised, на английском языке. Описание произведения, (предисловие) а так же отзывы посетителей доступны на портале библиотеки ЛибКат.

- Название:Handbook of Enology: Volume 1
- Автор:
- Жанр:
- Год:неизвестен
- ISBN:нет данных
- Рейтинг книги:4 / 5. Голосов: 1
-
Избранное:Добавить в избранное
- Отзывы:
-
Ваша оценка:
Handbook of Enology: Volume 1: краткое содержание, описание и аннотация
Предлагаем к чтению аннотацию, описание, краткое содержание или предисловие (зависит от того, что написал сам автор книги «Handbook of Enology: Volume 1»). Если вы не нашли необходимую информацию о книге — напишите в комментариях, мы постараемся отыскать её.
is thus the same. It aims to provide practitioners, winemakers, technicians and enology students with foundational knowledge and the most recent research results. This knowledge can be used to contribute to a better definition of the quality of grapes and wine, a greater understanding of chemical and microbiological parameters, with the aim of ensuring satisfactory fermentations and predicting the evolution of wines, an7thd better mastery of wine stabilization processes. As a result, the purpose of this publication is to guide readers in their thought processes with a view to preserving and optimizing the identity and taste of wine and its aging potential.
This third English edition of
, is an enhanced translation from the 7h French 2017 edition, and is published in print as individual themed volumes and as a two-volume set, describing aspects of winemaking using a detailed, scientific approach. The authors, who are highly-respected enologists, examine winemaking processes, theorizing what constitutes a perfect technique and the proper combination of components necessary to produce a quality vintage. They also illustrate methodologies of common problems, revealing the mechanism behind the disorder, thus enabling a diagnosis and solution.
Volume 1:
The Microbiology of
Wine and Vinifications
Coverage includes: Wine microbiology; Yeasts; Yeast metabolism; The conditions for the development of yeasts; Lactic acid bacteria, their metabolism and their development in wine; Acetic bacteria; The use of sulfur dioxide in the treatment of musts and wines; Products and processes acting in addition to sulfur dioxide; Winemaking; The grape and its maturation; Harvesting and processing of grapes after harvest; Vinification in red and white wine making.
The target audience includes advanced viticulture and enology students, professors and researchers, and practicing grape growers and vintners.
Handbook of Enology: Volume 1 — читать онлайн ознакомительный отрывок
Ниже представлен текст книги, разбитый по страницам. Система сохранения места последней прочитанной страницы, позволяет с удобством читать онлайн бесплатно книгу «Handbook of Enology: Volume 1», без необходимости каждый раз заново искать на чём Вы остановились. Поставьте закладку, и сможете в любой момент перейти на страницу, на которой закончили чтение.
Интервал:
Закладка:
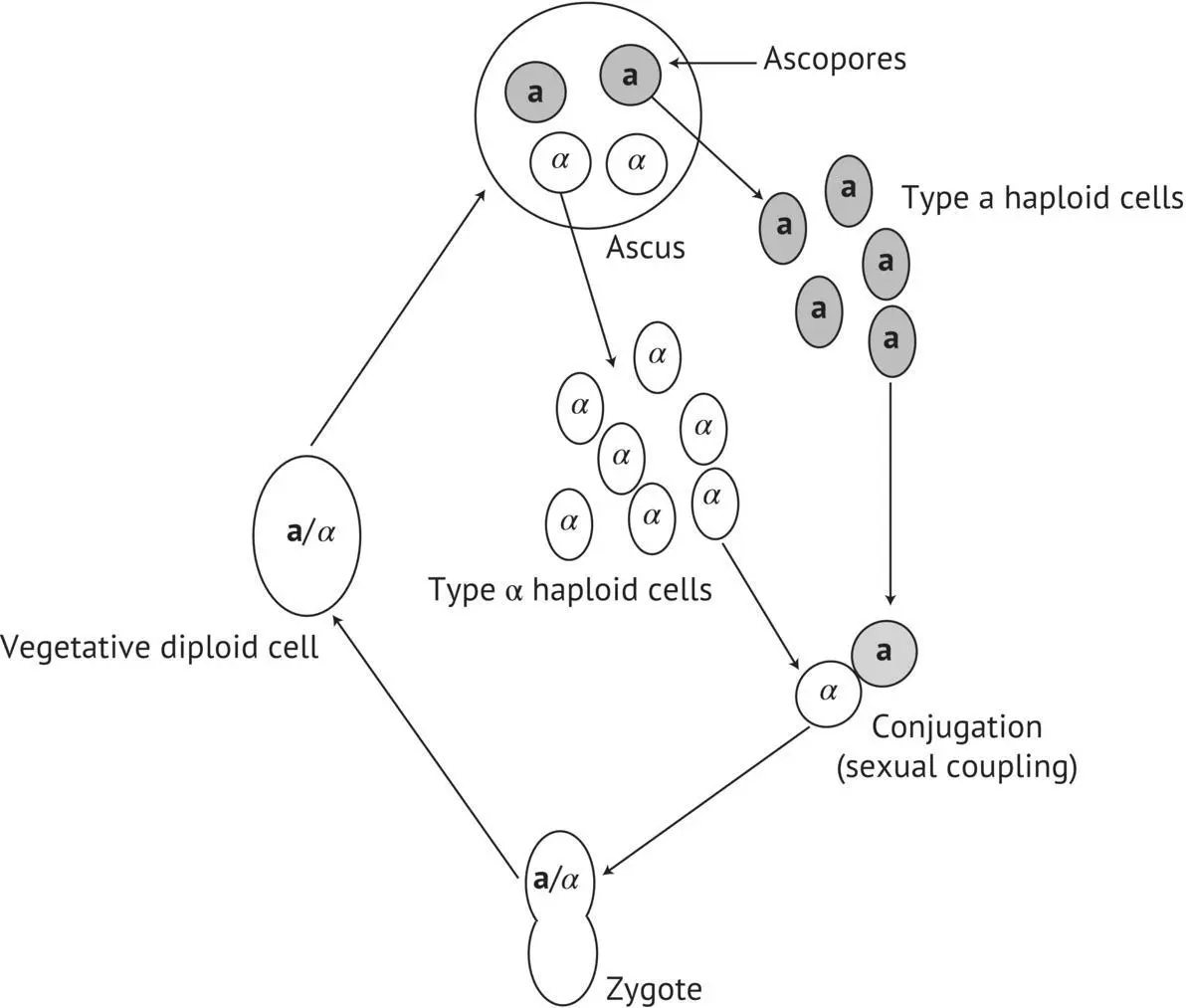
FIGURE 1.14 Reproduction cycle of a heterothallic yeast strain. a, α : spore mating types.
Most wild and selected winemaking strains that belong to the S. cerevisiae species are diploid and homothallic. This is also true of almost all of the strains that have been isolated in vineyards of the Bordeaux region. Moreover, recent studies carried out by Mortimer et al . (1994) in Californian and Italian vineyards have shown that the majority of strains (80%) are homozygous for the HO character ( HO / HO ); heterozygosis ( HO / ho ) is in the minority. Heterothallic strains ( ho / ho ) are rare (less than 10%). We have made the same observations for yeast strains isolated in the Bordeaux region; for example, the F10 strain, which is fairly prevalent in spontaneous fermentations in certain Bordeaux wines, is HO / HO . In other words, the four spores arising from an ascus give monoparental diploids, capable of forming asci when placed in a pure culture. This generalized homozygosis for the HO character of wild winemaking strains is probably an important factor in their evolution, according to the genome renewal phenomenon proposed by Mortimer et al . (1994) ( Figure 1.16). According to this author, the continuous reproduction of a yeast strain in its natural environment is accompanied by the accumulation of heterozygotic damage to the DNA. Certain slow‐growth or functional loss mutations of certain genes decrease strain vigor in the heterozygous state. Sporulation, however, produces haploid cells containing various combinations of these heterozygotic characters. All of these spores will become homozygous diploid cells with a series of genotypes because of the homozygosity of the HO character. Certain diploids that prove to be more vigorous than others will in time supplant the parents and less vigorous collaterals. This very persuasive model is supported by the characteristics of the wild winemaking strains analyzed. In these, the spore viability rate is an inverse function of the heterozygosis rate for a certain number of mutations. The completely homozygous strains present the highest spore viability and vigor.
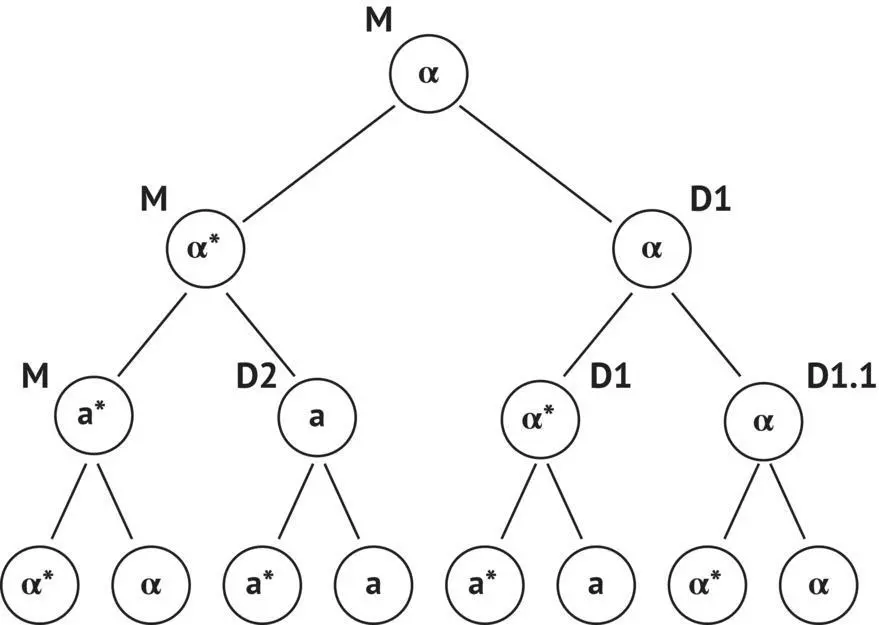
FIGURE 1.15 Mating type interconversion model of haploid yeast cells in a homothallic strain (Herskowitz et al ., 1992). * designates cells capable of changing mating type at the next cell division or cells already having undergone budding. M, initial cell carrying the HO gene; D1, D2, daughter cells of M; D1.1, daughter cell of D1.
In conclusion, we can question whether sporulation of strains under natural conditions is indispensable to ensure their growth and fermentation performance. We can also raise the question of conserving selected strains of active dry yeasts (ADYs) for use as yeast starters. It may be necessary to regenerate them periodically to eliminate possible mutations from their genome, which could diminish their vigor.
1.7 The Killer Phenomenon
1.7.1 Introduction
Certain yeast strains, known as killer strains (K), secrete protein toxins into their environment that are capable of killing other, sensitive strains (S). The killer strains are not sensitive to their own toxin but can be killed by a toxin that they do not produce. Additionally, neutral strains (N) do not produce a toxin but are resistant. The action of a killer strain on a sensitive strain is easy to demonstrate in the laboratory on an agar culture medium at pH 4.2–4.7 at 20°C. The sensitive strain is inoculated into the mass of agar before it solidifies; then the strain to be tested is inoculated in streaks on the solidified medium. If it is a killer strain, a clear zone in which the sensitive strain cannot grow encircles the inoculum streaks ( Figure 1.17).
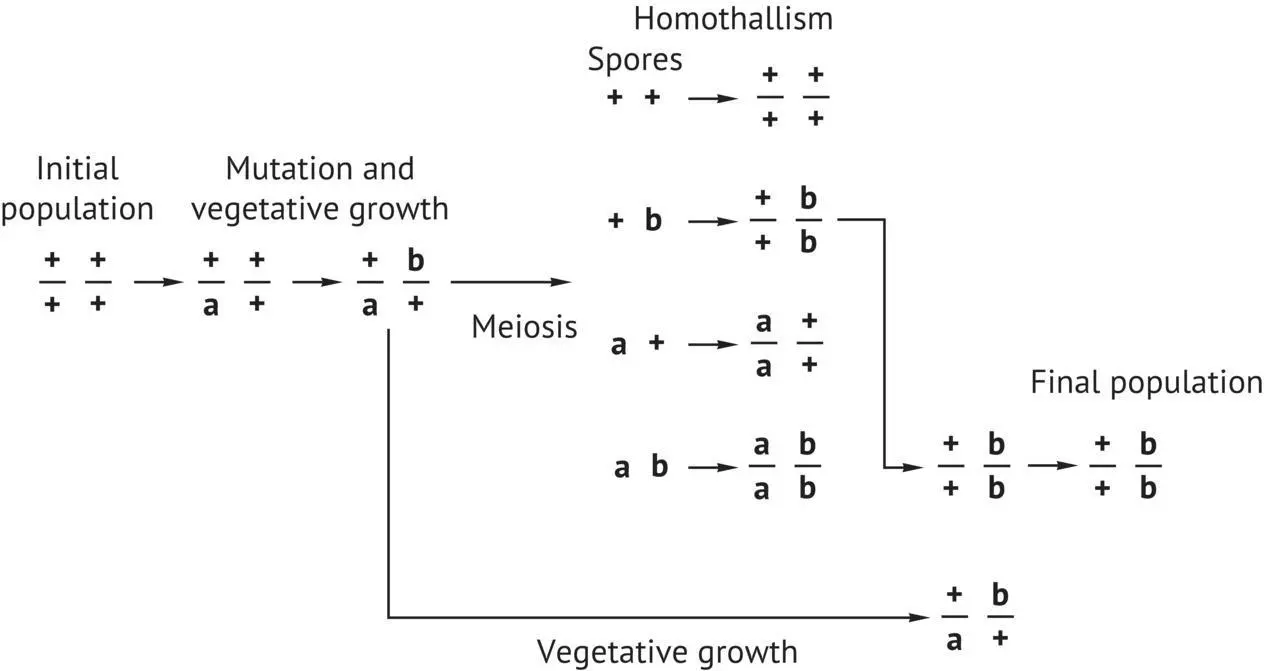
FIGURE 1.16 Genome renewal of a homozygote yeast strain for the HO gene of homothallism, having accumulated recessive mutations during vegetative reproduction (Mortimer et al ., 1994). (a and b), mutation of certain genes.
This phenomenon, known as the killer factor, was discovered in S. cerevisiae , but killer strains also exist in other yeast genera such as Hansenula , Candida , Kloeckera , Hanseniaspora , Pichia , Torulopsis , Kluyveromyces , and Debaryomyces . Killer yeasts have been classified into 11 groups according to the sensitivity reaction between strains as well as the nature and properties of the toxins involved. The killer factor is a cell interaction model mediated by the excreted protein toxin. It has given rise to much fundamental research (Tipper and Bostian, 1984; Young, 1987). Barre (1984, 1992), Radler (1988), and Van Vuuren and Jacobs (1992) have described the technological implications of this phenomenon for wine yeasts and the fermentation process.
1.7.2 Physiology and Genetics of the Killer Phenomenon
The determinants of the killer factor are both cytoplasmic and nuclear. In S. cerevisiae , the killer phenomenon is associated with the presence of double‐stranded RNA particles, called virus‐like particles (VLP), in the cytoplasm. They are in the same category as noninfectious mycoviruses. There are two kinds of VLP: M and L. The M genome (1.3–1.9 kb) codes for the toxin (K) and for the immunity factor (R). The L genome (4.5 kb) codes for an RNA polymerase and the protein capsid that encapsulates the two genomes. Killer strains (K +R +) secrete the toxin and are immune to it. The sensitive cells (K −R −) do not possess M VLP, but most of them have L VLP. The two types of viral particles are necessary for the yeast cell to express the killer phenotype (K +R +), since the L mycovirus is necessary for the maintenance of the M type.
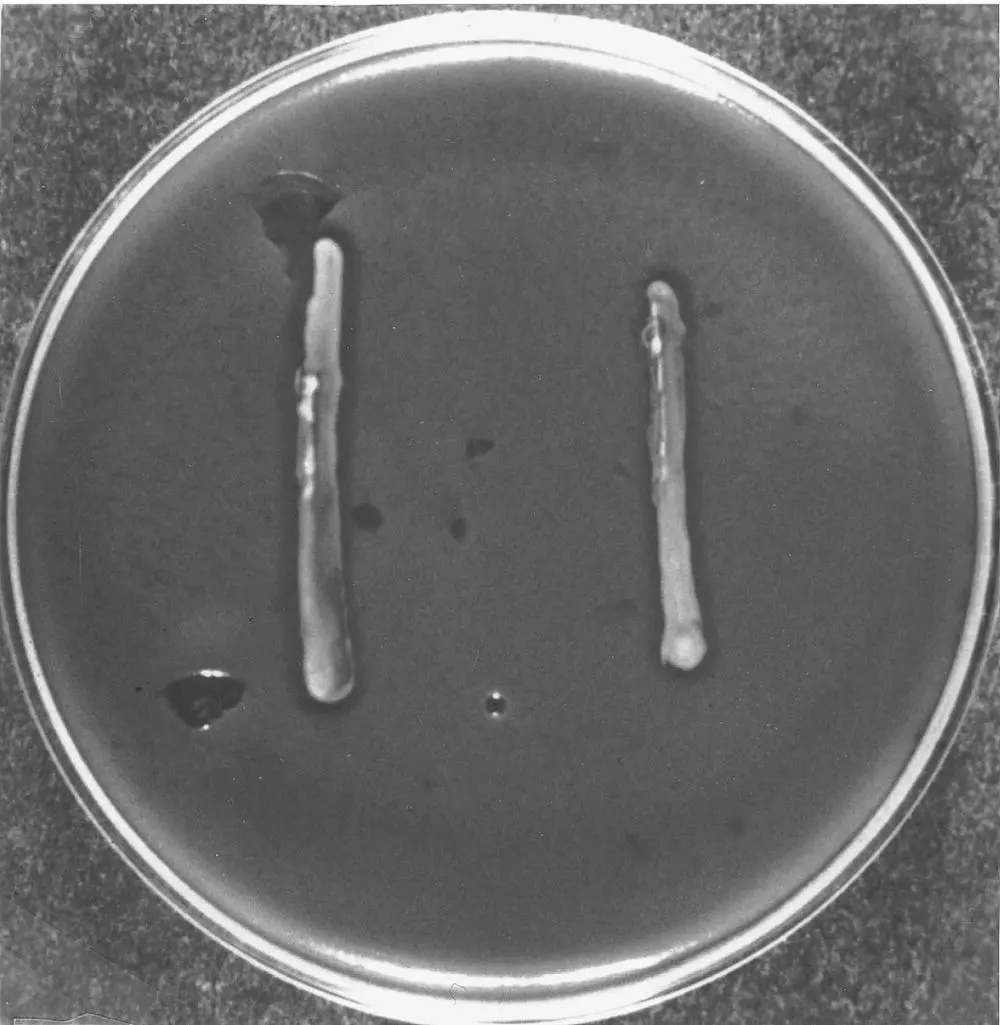
FIGURE 1.17 Identification of the K2 killer phenotype in S. cerevisiae . The presence of a halo around the two streaks of the killer strain is due to the death of the sensitive strain cultivated in the medium.
There are four types of killer activity in S. cerevisiae strains, corresponding to K1, K2, K28, and Klus toxins. They are, respectively, coded by M1, M2, M28, and Mlus VLPs with sizes of 1.8, 1.7, 2.1, and 2.3 kb. The K2 activity group is by far the most widespread in the S. cerevisiae strains encountered in wine (Rodríguez‐Cousiño et al ., 2011). Neutral strains (K −R +) are insensitive to a given toxin without being capable of producing it. They possess M VLPs of normal size that code only for the immunity factor, but they either do not produce toxins or are inactive because of mutations affecting the M‐type RNA.
Many chromosomic genes are involved in the maintenance and replication of L and M RNA particles as well as in the maturation and transport of the toxin produced.
The K1 toxin is a small protein made up of two subunits (9 and 9.5 kDa); it is active and stable in a very narrow pH range (4.2–4.6). It is therefore inactive in grape must. The K2 toxin, a 16 kDa glycoprotein produced by homothallic strains of S. cerevisiae encountered in wine, is active between pH 2.8 and 4.8 with a maximum activity between 4.2 and 4.4. It is therefore active at the pH of grape must and wine.
Читать дальшеИнтервал:
Закладка:
Похожие книги на «Handbook of Enology: Volume 1»
Представляем Вашему вниманию похожие книги на «Handbook of Enology: Volume 1» списком для выбора. Мы отобрали схожую по названию и смыслу литературу в надежде предоставить читателям больше вариантов отыскать новые, интересные, ещё непрочитанные произведения.












Обсуждение, отзывы о книге «Handbook of Enology: Volume 1» и просто собственные мнения читателей. Оставьте ваши комментарии, напишите, что Вы думаете о произведении, его смысле или главных героях. Укажите что конкретно понравилось, а что нет, и почему Вы так считаете.