Pascal Ribéreau-Gayon - Handbook of Enology - Volume 1
Здесь есть возможность читать онлайн «Pascal Ribéreau-Gayon - Handbook of Enology - Volume 1» — ознакомительный отрывок электронной книги совершенно бесплатно, а после прочтения отрывка купить полную версию. В некоторых случаях можно слушать аудио, скачать через торрент в формате fb2 и присутствует краткое содержание. Жанр: unrecognised, на английском языке. Описание произведения, (предисловие) а так же отзывы посетителей доступны на портале библиотеки ЛибКат.

- Название:Handbook of Enology: Volume 1
- Автор:
- Жанр:
- Год:неизвестен
- ISBN:нет данных
- Рейтинг книги:4 / 5. Голосов: 1
-
Избранное:Добавить в избранное
- Отзывы:
-
Ваша оценка:
Handbook of Enology: Volume 1: краткое содержание, описание и аннотация
Предлагаем к чтению аннотацию, описание, краткое содержание или предисловие (зависит от того, что написал сам автор книги «Handbook of Enology: Volume 1»). Если вы не нашли необходимую информацию о книге — напишите в комментариях, мы постараемся отыскать её.
is thus the same. It aims to provide practitioners, winemakers, technicians and enology students with foundational knowledge and the most recent research results. This knowledge can be used to contribute to a better definition of the quality of grapes and wine, a greater understanding of chemical and microbiological parameters, with the aim of ensuring satisfactory fermentations and predicting the evolution of wines, an7thd better mastery of wine stabilization processes. As a result, the purpose of this publication is to guide readers in their thought processes with a view to preserving and optimizing the identity and taste of wine and its aging potential.
This third English edition of
, is an enhanced translation from the 7h French 2017 edition, and is published in print as individual themed volumes and as a two-volume set, describing aspects of winemaking using a detailed, scientific approach. The authors, who are highly-respected enologists, examine winemaking processes, theorizing what constitutes a perfect technique and the proper combination of components necessary to produce a quality vintage. They also illustrate methodologies of common problems, revealing the mechanism behind the disorder, thus enabling a diagnosis and solution.
Volume 1:
The Microbiology of
Wine and Vinifications
Coverage includes: Wine microbiology; Yeasts; Yeast metabolism; The conditions for the development of yeasts; Lactic acid bacteria, their metabolism and their development in wine; Acetic bacteria; The use of sulfur dioxide in the treatment of musts and wines; Products and processes acting in addition to sulfur dioxide; Winemaking; The grape and its maturation; Harvesting and processing of grapes after harvest; Vinification in red and white wine making.
The target audience includes advanced viticulture and enology students, professors and researchers, and practicing grape growers and vintners.
Handbook of Enology: Volume 1 — читать онлайн ознакомительный отрывок
Ниже представлен текст книги, разбитый по страницам. Система сохранения места последней прочитанной страницы, позволяет с удобством читать онлайн бесплатно книгу «Handbook of Enology: Volume 1», без необходимости каждый раз заново искать на чём Вы остановились. Поставьте закладку, и сможете в любой момент перейти на страницу, на которой закончили чтение.
Интервал:
Закладка:
A single plasmid, called the 2 μm plasmid, has been identified in the yeast nucleus. It is a circular molecule of DNA, containing 6 kb, and there are 50–100 copies per cell. Its biological function is not known. However, it is a very useful tool, used by molecular biologists to construct artificial plasmids and genetically transform yeast strains.
1.6 Reproduction and the Yeast Biological Cycle
Like other spore‐forming yeasts belonging to the Ascomycetes class, S. cerevisiae can multiply either asexually by vegetative reproduction or sexually by forming ascospores. By definition, yeasts belonging to the imperfect fungi class can only reproduce by vegetative reproduction.
1.6.1 Vegetative Reproduction
Most yeasts undergo vegetative reproduction by a process called budding. Some yeasts, such as species belonging to the genus Schizosaccharomyces , reproduce by binary fission.
Figure 1.11(Tuite and Oliver (1991)) represents the life cycle of S. cerevisiae divided into four phases: M, G1, S, and G2. M corresponds to mitosis; G1 is the period preceding S, which is the synthesis of DNA; and G2 is the period before mitosis. As soon as the bud emerges, at the beginning of S, the splitting of the SPB can be observed in the nuclear membrane by electron microscopy, while the cytoplasmic microtubules orient themselves toward the emerging bud. These microtubules seem to guide numerous vesicles that appear in the budding zone and are involved in the reshaping of the cell wall. As the bud grows larger, discontinuous nuclear microtubules begin to appear. The longest microtubules form the mitotic spindle between the two SPBs. At the end of G2, the nucleus begins to push and pull apart in order to penetrate the bud. Some of the mitochondria also pass with some small vacuoles into the bud, whereas a large vacuole is formed at the other pole of the cell. The expansion of the latter seems to push the nucleus into the bud. During mitosis itself, the nucleus stretches to its maximum, and the mother cell separates from the daughter cell. This separation takes place only after the construction of the separation cell wall and the depositing of a ring of chitin on the bud scar of the mother cell. The movement of chromosomes during mitosis is difficult to observe in yeasts, but a microtubule–centromere link almost certainly guides the chromosomes. In grape must, the duration of budding is approximately one to two hours. As a result, the cell population doubles in two hours during the active yeast growth phase of fermentation.
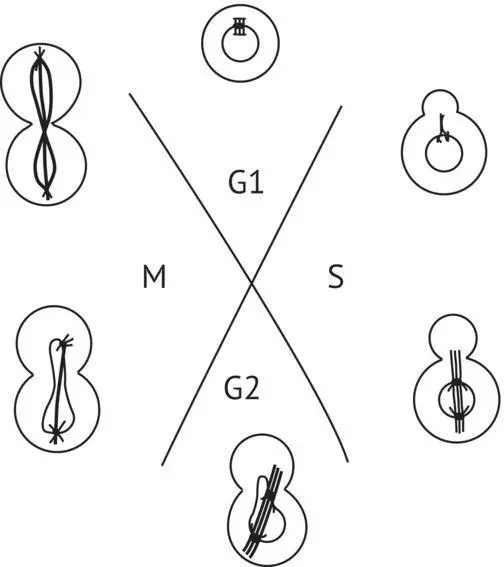
FIGURE 1.11 Saccharomyces cerevisiae cell cycle (vegetative reproduction) (Tuite and Oliver, 1991). M, mitosis; G1, period preceding DNA synthesis; S, DNA synthesis; G2, period preceding mitosis.
1.6.2 Sexual Reproduction
When the diploid cells of spore‐forming yeast are in a hostile nutrient medium (for example, depleted of fermentable sugar, poor in nitrogen, and very aerated), they stop multiplying. Some transform into a kind of sac with a thick cell wall. These sacs are called asci. Each one contains four haploid ascospores arising from meiotic division of the nucleus. Grape must and wine are not propitious to yeast sporulation and, in principle, it never occurs in this medium. Yet Mortimer et al . (1994) observed the sporulation of certain wine yeast strains, even in sugar‐rich media. Our researchers have often observed asci in “old” agar culture media stored for several weeks in the refrigerator or at ambient temperatures ( Figure 1.12). The natural conditions under which wild wine yeasts sporulate and the frequency of this phenomenon are not known. In the laboratory, the agar or liquid media conventionally used to provoke sporulation have a sodium acetate base (1%). In S. cerevisiae , aptitude for sporulation varies greatly from strain to strain. Wine yeasts, both wild and selected, do not sporulate easily, and when they do, they often produce nonviable spores.

FIGURE 1.12 Scanning electron microscope photograph of S. cerevisiae cells kept on a sugar‐agar medium for several weeks. Asci containing ascospores can be observed.
( Source: Photograph from M. Mercier, Department of Electron Microscopy, Université de Bordeaux 1.)
Meiosis in yeasts and in higher eukaryotes ( Figure 1.13) has some similarities. Several hours after the transfer of diploid vegetative cells to a sporulation medium, the SPB splits during the S phase of DNA replication. A dense body (DB) appears simultaneously in the nucleus near the nucleolus. The DB evolves into synaptonemal complexes—structures permitting the coupling and recombination of homologous chromosomes. After eight to nine hours in the sporulation medium, the two SPBs separate and the spindle begins to form. This stage is called metaphase I of meiosis. At this stage, the chromosomes are not yet visible. Then, while the nuclear membrane remains intact, the SPB divides. At metaphase II, a second mitotic spindle stretches itself while the ascospore cell wall begins to form. Spindle stretching and cell wall development of ascospores go hand in hand. Nuclear buds, cytoplasm, and organelles migrate into the ascospores. At this point, edification of the cell wall is completed. The spindle then disappears when the division is completed.
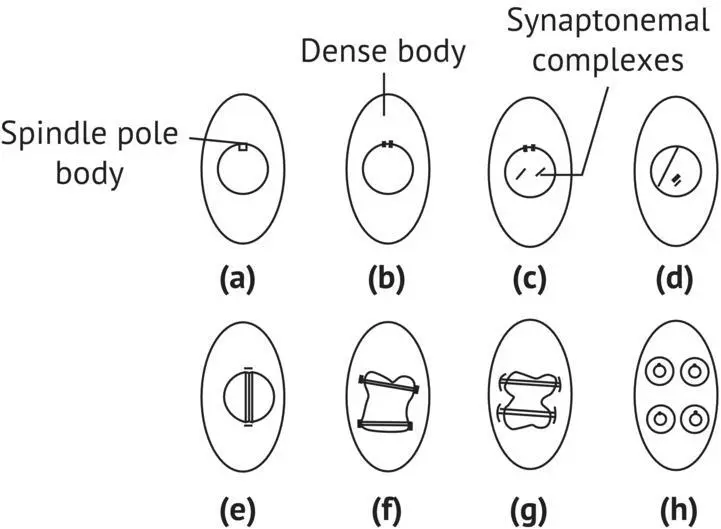
FIGURE 1.13 Meiosis in S. cerevisiae (Tuite and Oliver, 1991). (a) Cell before meiosis; (b) dividing of SPB; (c) synaptonemal complexes appear; (d) separation of SPBs; (e) constitution of spindle (metaphase I of meiosis); (f) dividing of the SPBs; (g) metaphase II of meiosis; (h) end of meiosis; formation of ascospores.
Placed under favorable conditions, i.e. sugar‐enriched nutrient media, the ascospores germinate, breaking the cell wall of the ascus, and begin to multiply. In S. cerevisiae , the haploid cells have two mating types: aand α . The ascus contains two aascospores and two α ascospores ( Figure 1.14). Type a( MAT a) cells produce a mating pheromone a. This peptide made up of 12 amino acids is called mating factor α . In the same manner, type α cells produce mating factor α , a peptide made up of 13 amino acids. The afactor, emitted by the MAT acells, stops the reproduction of MATα cells in G1, and reciprocally, the α factor produced by α cells stops the biological cycle of type acells. Mating occurs between two cells of the opposite mating type. Their agglutination enables cellular and nuclear fusion and makes use of cell wall glycoproteins, called aand α agglutinins. The vegetative diploid cell that is formed ( a/ α ) can no longer produce mating pheromones and is insensitive to their action; it reproduces by budding.
Some strains, from a monosporic culture, can be maintained in a stable haploid state. Their mating type remains constant for many generations. They are heterothallic. Others change mating type during cell division, causing diploid cells to appear in the descendants of an ascospore. They are homothallic and have an HO gene that inverses mating type at a high frequency during vegetative division. This interconversion ( Figure 1.15) occurs in the mother cell at the G1 stage of the biological cycle, after the first budding but before the DNA replication phase. In this manner, a type α M ascospore divides to produce two α cells (M and the first daughter cell, D1). During the following cell division, M produces two cells (M and D2) that have become acells. In the same manner, the D1 cell produces two α cells after the first division and two acells during its second budding. Laboratory strains that are deficient or missing the HO gene have a stable mating type. Heterothallism can therefore be considered the result of a mutation of the HO gene or of genes that control its functioning (Herskowitz et al ., 1992).
Читать дальшеИнтервал:
Закладка:
Похожие книги на «Handbook of Enology: Volume 1»
Представляем Вашему вниманию похожие книги на «Handbook of Enology: Volume 1» списком для выбора. Мы отобрали схожую по названию и смыслу литературу в надежде предоставить читателям больше вариантов отыскать новые, интересные, ещё непрочитанные произведения.












Обсуждение, отзывы о книге «Handbook of Enology: Volume 1» и просто собственные мнения читателей. Оставьте ваши комментарии, напишите, что Вы думаете о произведении, его смысле или главных героях. Укажите что конкретно понравилось, а что нет, и почему Вы так считаете.