Modern Trends in Structural and Solid Mechanics 3
Здесь есть возможность читать онлайн «Modern Trends in Structural and Solid Mechanics 3» — ознакомительный отрывок электронной книги совершенно бесплатно, а после прочтения отрывка купить полную версию. В некоторых случаях можно слушать аудио, скачать через торрент в формате fb2 и присутствует краткое содержание. Жанр: unrecognised, на английском языке. Описание произведения, (предисловие) а так же отзывы посетителей доступны на портале библиотеки ЛибКат.

- Название:Modern Trends in Structural and Solid Mechanics 3
- Автор:
- Жанр:
- Год:неизвестен
- ISBN:нет данных
- Рейтинг книги:4 / 5. Голосов: 1
-
Избранное:Добавить в избранное
- Отзывы:
-
Ваша оценка:
Modern Trends in Structural and Solid Mechanics 3: краткое содержание, описание и аннотация
Предлагаем к чтению аннотацию, описание, краткое содержание или предисловие (зависит от того, что написал сам автор книги «Modern Trends in Structural and Solid Mechanics 3»). Если вы не нашли необходимую информацию о книге — напишите в комментариях, мы постараемся отыскать её.
Modern Trends in Structural and Solid Mechanics 3 — читать онлайн ознакомительный отрывок
Ниже представлен текст книги, разбитый по страницам. Система сохранения места последней прочитанной страницы, позволяет с удобством читать онлайн бесплатно книгу «Modern Trends in Structural and Solid Mechanics 3», без необходимости каждый раз заново искать на чём Вы остановились. Поставьте закладку, и сможете в любой момент перейти на страницу, на которой закончили чтение.
Интервал:
Закладка:
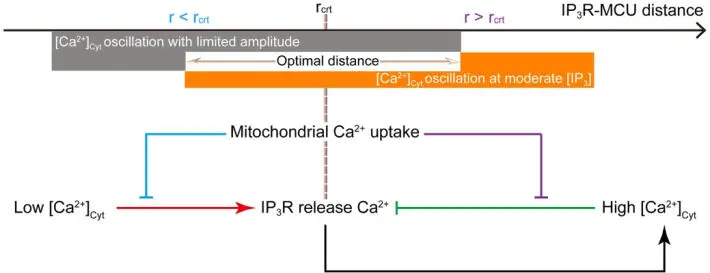
Figure 1.4. Schematic representation of how mitochondria modulate [Ca 2+] Cyt. Identifying the critical distance at which 50% of the IP 3R- released Ca 2+ ions are taken up by mitochondria. Outside this range, negative and positive control occurs on the Ca 2+ (Qi et al. (2015), with permission). For a color version of this figure, see www.iste.co.uk/challamel/mechanics3.zip
Figure 1.4 schematically represents an optimal microdomain distance for Ca 2+uptake. Figure 1.5(a) and (b) shows the Ca 2+fluctuations for cases with (a) and without (b) the presence of mitochondria.
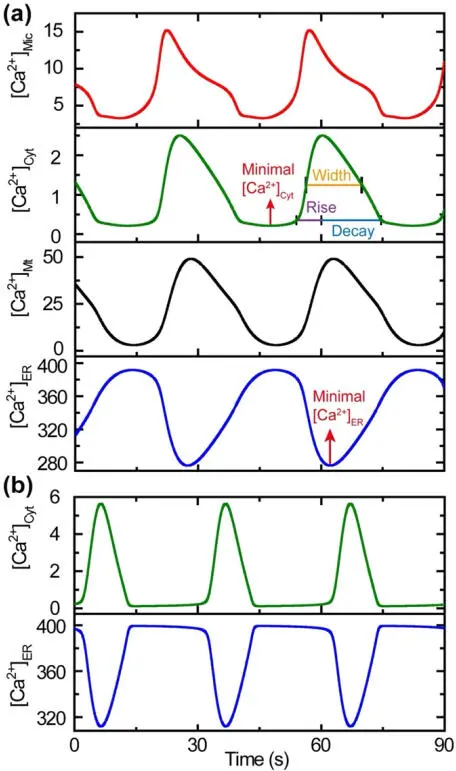
Figure 1.5. Mitochondria serve as Ca2+ reservoirs. The minimal values of [Ca2+]ER are 311 mM and 276 mM in the absence (b) and in the presence (a) of mitochondria, respectively, indicating that more Ca2+ ions are released from the ER during each spiking cycle in the presence of mitochondria. The maximal values of [Ca2+]Cyt are 5.6 mM and 2.5 mM in the absence (b) and in the presence of mitochondria (a), respectively, showing that mitochondria can significantly decrease [Ca2+]Cyt oscillation amplitude (Qi et al. (2015), with permission). For a color version of this figure, see www.iste.co.uk/challamel/mechanics3.zip
Mitochondria also have bidirectional communications with other cellular organelles, in particular, lysosomes (spherical vesicles that contain enzymes that can break down many kinds of biomolecules) and peroxisomes (organelles involved in the catabolism of various acids, in addition to other tasks). These communications are involved in the pathology of mitochondrial diseases (Diogo et al . 2018). These communications also provide coupling between the organelles and, while having evolved for efficiencies and survival, can also cause coupled dysfunctions. For example, lysosomes and peroxisomes are affected structurally and functionally by genetic defects in mitochondrial proteins that are known as mitochondrial diseases. Similarly, lysosomal and peroxisomal diseases perturb mitochondria. Lysosomal storage diseases perturb peroxisomal metabolism and mitochondrial function. Peroxisomal diseases often lead to alterations in mitochondrial structure, redox (ROS) balance and metabolism. The saturation of lysosomal capacity is often observed in mitochondrial diseases, with an accumulation of dysfunctional lysosomes and autophagosomes. Such couplings challenge both the modeling and the development of focused clinical treatments. Given all these couplings, we hypothesize that there can be a variety of optimal behavioral regimes that avoid pathological pathways. A goal can be to identify these via dynamic governing models.
In summary, an appropriate set of optimal interaction mechanisms need to be in place for healthy cell-wide functioning. In this context, a full understanding of cellular metabolism and mitochondrial diseases requires an understanding of mitochondrial communications with the rest of the cell, as well as within the organelle.
1.5. Mitochondrial motility
Mitochondrial motility, or trafficking, is critical for the survival of all cells. Neurons, which extend their axons and dendrites up to a meter, between two and three orders of magnitude greater than most other cells, are especially vulnerable to any inability by their mitochondria to get to sites with a high energy demand. At any instant of time, about 10–40% of the mitochondria are generally moving, with about half of those moving away from the cell body (anterograde, kinesin-dependent) and the rest towards the cell body (retrograde, dynein-dependent) (Schwarz 2013).
The distribution of mitochondria over long distances in the neurons is regulated by a complex molecular machinery that has evolved to match the very dynamic demand for energy with an optimal mitochondrial distribution (Schwarz 2013). Highly branched paths of the complex neuron geometry must be navigated, knowing where and when to stop. Machinery for fission and fusion can intersect with machinery for motility using feedforward and feedback mechanisms. Misregulation of motility can lead to neurodegeneration (Vanhauwaert et al . 2019).
The dendritic synapses are where the neurons receive signals from other neurons. Energy demand is greatest at the synapses and can change rapidly in response to almost instantaneous environmental changes. The same is true at the axonal synapses, which are used by neurons to transfer signaling downstream to subsequent neurons. Neuronal axons lie flat and are typically about a micron in diameter. Trafficking is along linear arrays of uniformly polarized microtubules, where the negative ends of the tubules are anchored in the cell body and the positive charge ends in the distal tips. Due to the local morphology, axonal mitochondria have separated from the reticulum and exist as discrete organelles of a dimension, typically 1–3 microns, with those in dendrites tending to be longer.
Mitochondria are a fundamental component for healthy living, supporting optimal functioning and efficient energy usage at all levels, thus avoiding numerous pathologies. In the discussion so far, we maintain that a significant controlling aspect of mitochondrial functioning is based on optimizations of a variety of defining characteristics. Here, motility and the placement of mitochondria within dendrites and axons can be viewed as optimal solutions, assuring sufficient energy at locations of high energy demand.
1.6. Cristae, ultrastructure and supercomplexes
Mitochondrial dynamics has evolved to define a broad array of actions and activities: already discussed are fission and fusion, and also cristae modifications, the constant changing of shape at the macro- and ultrastructural levels. Such modifications in shape are directly connected to their bioenergetic function. The cristae, where the respiratory chain complexes exist, have junctions of width between 20 nm and 40 nm. They control metabolite and protein access to the interior volume, known as the cristae lumen. They also control the apoptosis process of cell death. The junction structure and lumen are modified for more efficient respiration. So-called supercomplexes are created as part of the cristae remodeling, and these are required for optimal mitochondrial respiration. Slight morphological changes can fine-tune the efficiency of energy-producing respiration. There is a correlation between mitochondrial respiratory capacity and the number of cristae (Baker et al . 2019). A clear understanding of the supercomplexes, as well as the basis upon which cristae remodeling is optimized, can provide us with clues for clinical interventions that target respiratory-based diseases. We can hypothesize that morphological, mechanical and biochemical aspects play a role in cristae remodeling optimization, with the following discussion being of relevance.
The inner mitochondrial membrane and the cristae structure have only recently started to be understood. The inner membrane has the larger area even though it is enclosed within an outer membrane of a smaller area. To accommodate this, broad folds, called cristae, have evolved and project into the mitochondrial matrix where the bioenergetic processes occur. The observed morphology of the inner mitochondrial membrane is the result of the minimization of the system’s free energy. This free energy is a function of local bending energy and curvatures, the surface area, pressure differences and surface tension exerted by proteins. It appears, based on analytical models of these characterizing parameters, that the surface tension and tensile forces act as constraints on the morphology. The varied shapes of the cristae are defined by the stationary states of the above minimization. It appears that motor protein tensile forces are responsible for a stable inner membrane shape (Ghochani et al . 2010). The nanoscale cristae, regulated by molecular mechanisms, influence mitochondrial function (Mannella et al . 2013).
Читать дальшеИнтервал:
Закладка:
Похожие книги на «Modern Trends in Structural and Solid Mechanics 3»
Представляем Вашему вниманию похожие книги на «Modern Trends in Structural and Solid Mechanics 3» списком для выбора. Мы отобрали схожую по названию и смыслу литературу в надежде предоставить читателям больше вариантов отыскать новые, интересные, ещё непрочитанные произведения.












Обсуждение, отзывы о книге «Modern Trends in Structural and Solid Mechanics 3» и просто собственные мнения читателей. Оставьте ваши комментарии, напишите, что Вы думаете о произведении, его смысле или главных героях. Укажите что конкретно понравилось, а что нет, и почему Вы так считаете.