Николай Печуркин - Энергия и жизнь
Здесь есть возможность читать онлайн «Николай Печуркин - Энергия и жизнь» весь текст электронной книги совершенно бесплатно (целиком полную версию без сокращений). В некоторых случаях можно слушать аудио, скачать через торрент в формате fb2 и присутствует краткое содержание. Город: Новосибирск, Год выпуска: 1988, Издательство: Наука, сибирское отделение, Жанр: sci_biophys, на русском языке. Описание произведения, (предисловие) а так же отзывы посетителей доступны на портале библиотеки ЛибКат.

- Название:Энергия и жизнь
- Автор:
- Издательство:Наука, сибирское отделение
- Жанр:
- Год:1988
- Город:Новосибирск
- ISBN:нет данных
- Рейтинг книги:4 / 5. Голосов: 1
-
Избранное:Добавить в избранное
- Отзывы:
-
Ваша оценка:
Энергия и жизнь: краткое содержание, описание и аннотация
Предлагаем к чтению аннотацию, описание, краткое содержание или предисловие (зависит от того, что написал сам автор книги «Энергия и жизнь»). Если вы не нашли необходимую информацию о книге — напишите в комментариях, мы постараемся отыскать её.
Книга будет интересна для биологов, физиков, химиков, биофизиков, а также всех интересующихся общими вопросами развития.
Энергия и жизнь — читать онлайн бесплатно полную книгу (весь текст) целиком
Ниже представлен текст книги, разбитый по страницам. Система сохранения места последней прочитанной страницы, позволяет с удобством читать онлайн бесплатно книгу «Энергия и жизнь», без необходимости каждый раз заново искать на чём Вы остановились. Поставьте закладку, и сможете в любой момент перейти на страницу, на которой закончили чтение.
Интервал:
Закладка:
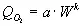 ,
,
где а и k — константы.
«Справедливость этого уравнения установлена для большинства групп животных, от простейших до млекопитающих. Можно сказать, что эта зависимость является эмпирическим законом, справедливым для всех животных», [3отин, Зотина, 1976, с. 49]. Величина k , меньшая единицы, хорошо соответствует известному закону поверхностей Рубнера, так как метаболизм соответствует массе, числу функционирующих клеток, а теплоотдача идет с поверхности. Это означает, что с ростом размеров организма падает величина удельной теплопродукции, так как уменьшается отношение поверхность: объем. Для нашего анализа пока больший интерес представляют данные о том, что в процессе эволюции, а не индивидуального развития коэффициент a , характеризующий в данном уравнении интенсивность энергообмена, существенно возрастает. Согласно данным, суммированным Хеммингсеном для трех, далеко отстоящих по организации групп организмов, константы уравнения равны в среднем: для одноклеточных a 1= 0,084 кал/ч; для пойкилотермных a 2= 0,69; для гомойотермных животных a 3= 19,68 кал/ч. Следовательно, интенсивность обмена возрастает от простейших к гомойотермным более чем на два порядка, более чем в 200 раз. Поддержание постоянной температуры тела для гомойотермных животных обходится примерно в 30 раз дороже по тратам на обмен, по сравнению с пойкилотермными того же размера. Казалось бы, огромная расточительность! Однако преимущества такого ароморфоза позволили гомойотермным животным занять места, недоступные для пойкилотермов. Вспомним белых медведей и песцов в Арктике или пингвинов в Антарктике; очень впечатляют с этой точки зрения сезонные миграции птиц на многие тысячи километров. Пример с птицами особенно наглядно показывает резко возросшие энергетические возможности гомойотермных животных.
К сожалению, данных по полному энергетическому обмену организмов, тем более для популяционпого или экосистемвого уровня, явно недостаточно. Приведенные выше результаты относятся главным образом к основному энергетическому обмену, который составляет лишь часть полного. Хотя наиболее вероятно их параллельное возрастание, так как за каждое новое «изобретение» организму необходимо расплачиваться прежде всего дополнительным расходом энергии. Но в целом возрастание активного обмена и есть итоговая мера прогресса.
Особенно наглядны расхождения обменов для человеческой популяции, вовлекающей в свою среду дополнительные энергетические источники. Если по уровню основного обмена человек занимает срединное положение в группе млекопитающих (согласно известной диаграмме «от мыши до слона»), то дополнительное использование и производство энергоресурсов у него в среднем в 20 раз выше. А в развитых странах и в 50–100 раз! Следовательно, согласно ЭПИР, человек в 20 раз более активен, чем млекопитающие и птицы, обладающие самыми высокими показателями энергорассеяния.
Б. Рост размеров особи и длительности поколения.В теории морфофизиологической эволюции рост продолжительности жизни и уменьшение числа потомков являются одним из показателей увеличения приспособленности животных. Для всего развития жизни в условиях нехватки вещества и постоянной накачки энергией это почти очевидно: резко снижается зависимость от лимитирования по веществу и сохраняется возможность использовать энергетические потоки.
С виду несколько противоречащим ЭПИР кажется увеличение средних размеров организмов в эволюции. Однако не следует забывать, что с увеличением размера организмов, согласно уравнению теплообмена, падает основной обмен, в то время как активный обмен возрастает. (Например, более крупные животные способны перемещаться на большие расстояния и из большего числа выбирать места расселения.)
Подчеркнем высокую напряженность обмена у мелких животных, т. е. большую долю поддерживающего метаболизма по сравнению с активным. Подсчитано, например, что маленькие грызуны расходуют до 95% своей энергии на поддержание постоянной температуры тела и основной метаболизм — тут переваривание и усвоение пищи, работа внутренних органов, мышц, нервной системы — и лишь единицы процентов на активную жизнь: миграции, поиски полового партнера, освоение новых местообитаний [Шварц, 1980]. Относительная доля поддерживающих трат у мелких млекопитающих в десятки раз больше, чем у крупных.
Читать дальшеИнтервал:
Закладка:
Похожие книги на «Энергия и жизнь»
Представляем Вашему вниманию похожие книги на «Энергия и жизнь» списком для выбора. Мы отобрали схожую по названию и смыслу литературу в надежде предоставить читателям больше вариантов отыскать новые, интересные, ещё непрочитанные произведения.












Обсуждение, отзывы о книге «Энергия и жизнь» и просто собственные мнения читателей. Оставьте ваши комментарии, напишите, что Вы думаете о произведении, его смысле или главных героях. Укажите что конкретно понравилось, а что нет, и почему Вы так считаете.