Михаил Никитин - Происхождение жизни. От туманности до клетки
Здесь есть возможность читать онлайн «Михаил Никитин - Происхождение жизни. От туманности до клетки» — ознакомительный отрывок электронной книги совершенно бесплатно, а после прочтения отрывка купить полную версию. В некоторых случаях можно слушать аудио, скачать через торрент в формате fb2 и присутствует краткое содержание. Город: Москва, Год выпуска: 2016, ISBN: 2016, Издательство: Литагент Альпина, Жанр: Прочая научная литература, Биология, Химия, на русском языке. Описание произведения, (предисловие) а так же отзывы посетителей доступны на портале библиотеки ЛибКат.

- Название:Происхождение жизни. От туманности до клетки
- Автор:
- Издательство:Литагент Альпина
- Жанр:
- Год:2016
- Город:Москва
- ISBN:978-5-9614-4350-9
- Рейтинг книги:5 / 5. Голосов: 2
-
Избранное:Добавить в избранное
- Отзывы:
-
Ваша оценка:
Происхождение жизни. От туманности до клетки: краткое содержание, описание и аннотация
Предлагаем к чтению аннотацию, описание, краткое содержание или предисловие (зависит от того, что написал сам автор книги «Происхождение жизни. От туманности до клетки»). Если вы не нашли необходимую информацию о книге — напишите в комментариях, мы постараемся отыскать её.
Происхождение жизни. От туманности до клетки — читать онлайн ознакомительный отрывок
Ниже представлен текст книги, разбитый по страницам. Система сохранения места последней прочитанной страницы, позволяет с удобством читать онлайн бесплатно книгу «Происхождение жизни. От туманности до клетки», без необходимости каждый раз заново искать на чём Вы остановились. Поставьте закладку, и сможете в любой момент перейти на страницу, на которой закончили чтение.
Интервал:
Закладка:
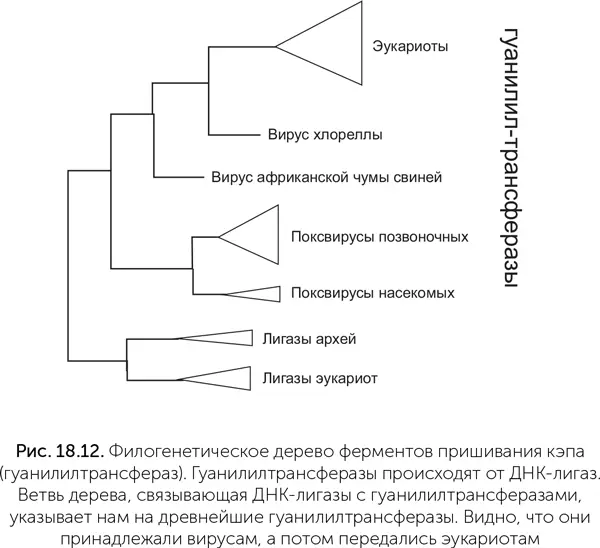
Чтобы понять, какой из узлов дерева гуанилилтрансфераз самый древний и соответствует предковому ферменту, к ним добавлены родственные ферменты – АТФ-зависимые ДНК-лигазы. Это более древние белки, предковые по отношению к гуанилилтрансферазам. Соответственно, та часть дерева гуанилилтрансфераз, которая выходит из лигазной части дерева, укажет нам древнейшую гуанилилтрансферазу. Оказывается, она принадлежала вирусам: первая ветвь ее потомков содержит ферменты поксвирусов (вирус оспы и его родственники), вторая – ферменты вируса ASF (африканской чумы свиней – African swine fever) и третья – вируса хлореллы. Гуанилилтрансферазы эукариот происходят от фермента вирусной линии, давшей начало вирусам ASF и хлореллы. Это доказывает, что кэпирование мРНК, как и другие инновации в генетических системах, появилось сначала среди вирусов и лишь затем было заимствовано эукариотами.
Для вирусов, в отличие от клеток, есть очевидная выгода в создании такой системы. Это средство перехвата управления клеткой хозяина. Вирус вносит в клетку фермент, который разрушает клеточные матричные РНК без кэпа и не трогает вирусные мРНК с кэпом. Белок eIF4E тоже мог сначала появиться у вирусов как средство захвата рибосом хозяина. Он присоединяется к рибосомам, после чего они узнают только вирусные мРНК с кэпом, останавливая синтез белков клетки. Современные клеточные eIF4E обычно связаны с рибосомами. У мимивируса [21]обнаружен собственный белок-аналог eIF4E, роль его пока непонятна.
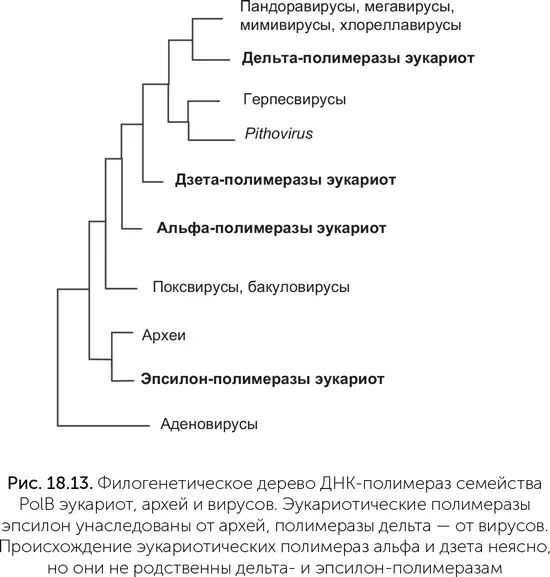
Такемура обратил внимание на систему репликации ДНК эукариот. По сравнению с машинами репликации бактерий и архей эукариотическая версия сложна и медлительна. Если у бактерий и архей скорость репликации составляет около 1000 нуклеотидов в секунду, то у эукариот обычно около 50. Там, где многие бактерии и археи обходятся одной ДНК-полимеразой (см. главу 14), эукариоты используют три родственные. Дельта-полимераза копирует отстающую цепь, эпсилон-полимераза – лидирующую, а альфа-полимераза имеет неожиданную функцию. Она удлиняет РНК-затравки на 15–20 нуклеотидов уже из ДНК, после чего уступает место полимеразам дельта и эпсилон. Альфа-полимераза часто делает ошибки, поэтому созданные ею фрагменты ДНК потом удаляются одновременно с РНК-затравками, и дельта-полимераза строит на их месте точные копии ДНК-матрицы. Кроме этих трех ДНК-полимераз у большинства эукариот есть четвертая – дзета-полимераза. Она заменяет другие полимеразы, когда нужно срочно скопировать поврежденную ДНК и нет времени на ее починку.
Все четыре эукариотические полимеразы принадлежат к семейству PolB (глава 14), но достаточно сильно различаются между собой. На родословном дереве ДНК-полимераз эукариот, архей и вирусов (рис. 18.13) хорошо видно, что полимераза эпсилон унаследована эукариотами от архей, полимераза дельта – от вирусов, а полимеразы альфа и дзета образуют отдельные ветви, начинающиеся ближе к корню дерева, чем δ. Об их происхождении нельзя судить точно, но они тоже могли попасть в клетки эукариот от каких-то вымерших или еще не открытых вирусов.
Наконец, Патрик Фортерр связал с вирусами происхождение огромного количества эукариотических белков, которые не похожи ни на какие белки бактерий и архей (Forterre, 2011). По последним оценкам, общий предок современных линий эукариот уже имел более 1400 таких белков. Среди них преобладают вспомогательные белки цитоскелета и ДНК-связывающие белки, в структуре которых присутствуют многочисленные короткие повторы. Вирусы, особенно крупные, содержат много генов, не похожих ни на какие гены клеточных организмов, а часто и на гены других вирусов. Кодируемые ими белки тоже часто имеют множественные короткие повторы. Низкая точность вирусных полимераз, интенсивная рекомбинация и «гонка вооружений» с хозяевами приводят к очень быстрой эволюции вирусных белков по сравнению с клеточными, поэтому вирусы – обильный источник принципиально новых белков.
Вирусная теория происхождения ядра и полового размножения
К крупным ДНК-вирусам (LNCDV, Large Nucleo-Cytoplasmic DNA Viruses) относятся, например, поксвирусы (из них широко известен возбудитель оспы), мимивирусы (недавно открытая группа гигантских вирусов-паразитов амеб) и фикоднавирусы, поражающие водоросли. Они имеют крупную и сложную вирусную частицу, покрытую несколькими мембранами. Самая внешняя мембрана сливается с наружной мембраной клетки при заражении, и в цитоплазму попадает вирус, окруженный двумя внутренними мембранами. Эти мембраны местами сливаются между собой, образуя некое подобие ядерных пор. Они выстланы изнутри белками капсида (вирусной оболочки), аналогичными белкам ядерной оболочки. В такой форме вирус долго существует в цитоплазме, в нем происходят транскрипция, кэпирование и полиаденилирование РНК, экспорт зрелых РНК в цитоплазму через поры вирусной частицы. Вирусы оспы раньше называли «мини-ядрами», а вирион мимивируса по размеру почти не уступает ядру клетки-хозяина.
Читать дальшеИнтервал:
Закладка:
Похожие книги на «Происхождение жизни. От туманности до клетки»
Представляем Вашему вниманию похожие книги на «Происхождение жизни. От туманности до клетки» списком для выбора. Мы отобрали схожую по названию и смыслу литературу в надежде предоставить читателям больше вариантов отыскать новые, интересные, ещё непрочитанные произведения.





![Михаил Никитин - Книга третья - Эпоха Барсума [СИ]](/books/428418/mihail-nikitin-kniga-tretya-epoha-barsuma-si-thumb.webp)




Обсуждение, отзывы о книге «Происхождение жизни. От туманности до клетки» и просто собственные мнения читателей. Оставьте ваши комментарии, напишите, что Вы думаете о произведении, его смысле или главных героях. Укажите что конкретно понравилось, а что нет, и почему Вы так считаете.