Михаил Никитин - Происхождение жизни. От туманности до клетки
Здесь есть возможность читать онлайн «Михаил Никитин - Происхождение жизни. От туманности до клетки» — ознакомительный отрывок электронной книги совершенно бесплатно, а после прочтения отрывка купить полную версию. В некоторых случаях можно слушать аудио, скачать через торрент в формате fb2 и присутствует краткое содержание. Город: Москва, Год выпуска: 2016, ISBN: 2016, Издательство: Литагент Альпина, Жанр: Прочая научная литература, Биология, Химия, на русском языке. Описание произведения, (предисловие) а так же отзывы посетителей доступны на портале библиотеки ЛибКат.

- Название:Происхождение жизни. От туманности до клетки
- Автор:
- Издательство:Литагент Альпина
- Жанр:
- Год:2016
- Город:Москва
- ISBN:978-5-9614-4350-9
- Рейтинг книги:5 / 5. Голосов: 2
-
Избранное:Добавить в избранное
- Отзывы:
-
Ваша оценка:
Происхождение жизни. От туманности до клетки: краткое содержание, описание и аннотация
Предлагаем к чтению аннотацию, описание, краткое содержание или предисловие (зависит от того, что написал сам автор книги «Происхождение жизни. От туманности до клетки»). Если вы не нашли необходимую информацию о книге — напишите в комментариях, мы постараемся отыскать её.
Происхождение жизни. От туманности до клетки — читать онлайн ознакомительный отрывок
Ниже представлен текст книги, разбитый по страницам. Система сохранения места последней прочитанной страницы, позволяет с удобством читать онлайн бесплатно книгу «Происхождение жизни. От туманности до клетки», без необходимости каждый раз заново искать на чём Вы остановились. Поставьте закладку, и сможете в любой момент перейти на страницу, на которой закончили чтение.
Интервал:
Закладка:
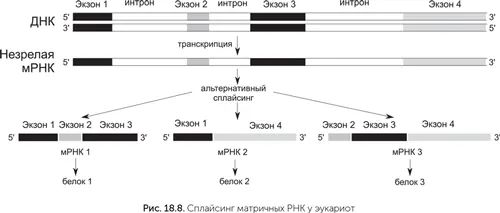
Не очень понятно, зачем эукариотам эти сложности. Сплайсинг дает некоторые преимущества, например, позволяет эукариотам получать с одного гена много разных белков. Для этого часть экзонов вырезается из мРНК вместе с интронами. В зависимости от того, какие экзоны войдут в зрелую матричную РНК, получатся разные варианты белка. Рекорд разнообразия альтернативного сплайсинга принадлежит белку иммунной системы насекомых DSCAM. Ген DSCAM мухи содержит 117 экзонов, комбинации которых дают 38 000 вариантов белка.
Эукариоты платят за эту возможность дополнительными затратами энергии и времени на синтез интронов мРНК. Хуже того, из-за сплайсинга включение и выключение генов занимает гораздо больше времени. У бактерий синтез белков на новой мРНК начинается еще до того, как она будет достроена до конца, благодаря этому через несколько минут после включения гена с него получатся первые готовые белки. У эукариот же сначала мРНК должна быть достроена до конца, а из-за интронов она длиннее, чем у бактерий, и синтезируется дольше. Потом должен пройти сплайсинг, тоже занимающий время, затем прикрепление кэпа, полиаденинового хвоста и экспорт мРНК из ядра в цитоплазму. Только после этого рибосомы смогут приступить к работе. Поэтому у эукариот от включения гена до появления первого готового белка проходит в лучшем случае от 30 до 60 минут, а часто несколько часов.
В ходе сплайсинга нередко происходят ошибки, приводящие к появлению дефектных мРНК. Для уничтожения таких матричных РНК эукариоты имеют специальную систему NMD (nonsence-mediated decay). Немногочисленные интроны в геномах бактерий вырезают себя из РНК сами, т. е. они являются рибозимами. В геномах эукариот счет интронов идет на десятки тысяч, и способностью к вырезанию самих себя они не обладают. Сплайсинг эукариот происходит при помощи специального РНК-белкового комплекса – сплайсосомы. Основную роль в ней играют шесть видов малых ядерных РНК, которые являются рибозимами. В структуре этих РНК есть сходство с самовырезающимися интронами бактерий.
Появление ядра, разделившее синтез матричных РНК и синтез белков, сняло эти ограничения, и началось бурное размножение интронов в белок-кодирующих генах предка эукариот. Мутации интронов часто нарушали их способность к самовырезанию и приводили к возникновению большого количества дефектных белков. Это вызвало давление отбора на появление системы NMD для контроля качества мРНК. Часть интронов специализировалась на вырезании других, поврежденных интронов. От них произошли малые ядерные РНК, составляющие основу сплайсосомы.
Гипотезы о происхождении ядра
Если происхождение митохондрий в целом понятно, то о пути появления ядра общепринятой теории нет. Образование ядра должно было в первую очередь обозначиться появлением оболочки вокруг генетического материала. Существует четыре гипотезы, объясняющие появление ядерной оболочки:
• гипотеза впячивания клеточной мембраны;
• эндоспоровая гипотеза;
• симбиотическая гипотеза;
• гипотеза слияния клеточных выростов.
Согласно гипотезе впячивания клеточной мембраны, оболочка ядра возникла из клеточной мембраны, часть которой ввернулась внутрь клетки. Так получилась система мембран эндоплазматического ретикулума, которая является продолжением двойной ядерной мембраны. Окружение ДНК мембраной было выгодно при питании путем фагоцитоза, так как при этом в цитоплазму неизбежно попадала чужеродная ДНК. Хуже того, в ней содержались вирусы и мобильные элементы. Поэтому появление ядерной оболочки для защиты генетического материала было поддержано отбором.
Подобное вворачивание мембраны и окружение ею ДНК известно у бактерии Gemmata obscuriglobis из группы Planctomycetes (Fuerst, Sagulenko, 2012) (рис. 18.9). Планктомицеты имеют наряду с наружной клеточной мембраной еще внутреннюю, которая делит цитоплазму клетки на две части. Внутренняя цитоплазма (пиреллюлосома) содержит рибосомы, и в ней происходит синтез белка, а внешняя цитоплазма (парифоплазма) не имеет рибосом. У Gemmata obscuriglobis внутренняя мембрана образует складку, которая окружает ДНК аналогично ядерной мембране эукариот. Однако у Gemmata эта оболочка не отделяет геномную ДНК от рибосом. Как и у всех бактерий, рибосомы Gemmata связываются с еще не завершенными матричными РНК и сразу начинают синтез белка. Ядерная мембрана эукариот же разделяет процессы транскрипции (создания матричных РНК) и трансляции (синтез белка по инструкциям в матричных РНК).
Читать дальшеИнтервал:
Закладка:
Похожие книги на «Происхождение жизни. От туманности до клетки»
Представляем Вашему вниманию похожие книги на «Происхождение жизни. От туманности до клетки» списком для выбора. Мы отобрали схожую по названию и смыслу литературу в надежде предоставить читателям больше вариантов отыскать новые, интересные, ещё непрочитанные произведения.





![Михаил Никитин - Книга третья - Эпоха Барсума [СИ]](/books/428418/mihail-nikitin-kniga-tretya-epoha-barsuma-si-thumb.webp)




Обсуждение, отзывы о книге «Происхождение жизни. От туманности до клетки» и просто собственные мнения читателей. Оставьте ваши комментарии, напишите, что Вы думаете о произведении, его смысле или главных героях. Укажите что конкретно понравилось, а что нет, и почему Вы так считаете.