Е. Бессолицына - Биохимия метаболизма. Учебное пособие
Здесь есть возможность читать онлайн «Е. Бессолицына - Биохимия метаболизма. Учебное пособие» — ознакомительный отрывок электронной книги совершенно бесплатно, а после прочтения отрывка купить полную версию. В некоторых случаях можно слушать аудио, скачать через торрент в формате fb2 и присутствует краткое содержание. ISBN: , Издательство: Литагент Ридеро, Жанр: Прочая научная литература, Биология, на русском языке. Описание произведения, (предисловие) а так же отзывы посетителей доступны на портале библиотеки ЛибКат.

- Название:Биохимия метаболизма. Учебное пособие
- Автор:
- Издательство:Литагент Ридеро
- Жанр:
- Год:неизвестен
- ISBN:9785448336638
- Рейтинг книги:3 / 5. Голосов: 1
-
Избранное:Добавить в избранное
- Отзывы:
-
Ваша оценка:
Биохимия метаболизма. Учебное пособие: краткое содержание, описание и аннотация
Предлагаем к чтению аннотацию, описание, краткое содержание или предисловие (зависит от того, что написал сам автор книги «Биохимия метаболизма. Учебное пособие»). Если вы не нашли необходимую информацию о книге — напишите в комментариях, мы постараемся отыскать её.
Биохимия метаболизма. Учебное пособие — читать онлайн ознакомительный отрывок
Ниже представлен текст книги, разбитый по страницам. Система сохранения места последней прочитанной страницы, позволяет с удобством читать онлайн бесплатно книгу «Биохимия метаболизма. Учебное пособие», без необходимости каждый раз заново искать на чём Вы остановились. Поставьте закладку, и сможете в любой момент перейти на страницу, на которой закончили чтение.
Интервал:
Закладка:
Четвертая стадия – превращение 3-фосфоглицерата в пируват
3-фосфоглицерат, изомеризуется в 2-фосфоглицерат под действием 3—2 фосфоглицерат изомеразы. 2-фосфоглицерат дегидратируется енолазой (2-фосфоглицератгидролиазой), происходит отщепление молекулы воды образуется фосфоенолпируват (ФЕП), который является «макроэргическим» соединением, фосфорильная группа которого может быть легко перенесена на AДФ (под действием фермента пируваткиназы); остающийся при этом енол пировиноградной кислоты самопроизвольно превращается в значительно более устойчивый пируват. Поскольку на каждую молекулу глюкозы образуются две молекулы фосфоенолпирувата, этот процесс восполняет затрату двух молекул АТФ, происходящую на начальных стадиях образования фруктозо-1,6-дифосфата из глюкозы.
Суммарная реакция гликолиза:
Глюкоза +2АТФ +2NAD +4АДФ = 2Пируват +2АДФ +4АТФ +2NADH
Суммарный энергетический выход всего 2 АТФ.
Энергетическая выгода гликолиза не велика, и используется организмами либо имеющими доступ большим количествам субстратов (моносахаридов), например паразитические организмы, либо из-за условий среды (анаэробные организмы).
Гликолиз – это один из древнейших метаболитических путей, по некоторым данным считается первые живые организмы получали энергию путем гликолиза. Поэтому механизмы регуляции гликолиза очень хорошо отрегулированы и направлены на обеспечение клетки энергией с одной стороны, но при этом на сохранение ресурсов клетки с другой. Механизмы регуляции гликолиза изучены достаточно хорошо.
Регуляция гликолиза
Регуляция гликолиза происходит на трех этапах:
Вход глюкозы в гликолиз (это естественно, так как если процесс не нужен, то его проще не запускать вообще, а не обрывать на половине).
Фосфофофруктокиназная реакция (реакция необратима, кроме того в ней затрачивается АТФ).
Пируваткиназная реакция (реакция также необратима, а кроме того важным является процесс утилизации образующегося пирувата).
Теперь необходимо рассмотреть эти этапы более подробно.
Вход глюкозы в гликолиз
Как было рассмотрено выше, глюкоза входит в гликолиз из свободной глюкозы или из гликогена. Свободная глюкоза фосфорилируется гексокиназой, активность этого фермента регулируется: происходит ингибирование продуктом реакции глюкозо-6-фосфатом. Поэтому накопление глюкозо-6-фосфата резко снижает скорость гексокиназной реакции, в результате нет затрат АТФ, так как глюкозо-6-фосфат во всех тканях кроме печени направляется на реакции окисления. Тогда как в печени накопление глюкозо-6-фосфата не происходит, так как излишки запасаются в виде гликогена, поэтому в печени работает другой фермент – глюкокиназа, не ингибируемый продуктом реакции.
При входе глюкозы из гликогена первая реакция гликогенфосфорилазная, регуляция гликогенфосфорилазы происходит двумя путями. Первый вариант посттрансляционная модификация.
В скелетных мышцах этот фермент присутствует в двух формах – в каталитически активной фосфорилированной форме (фосфорилаза а) и в значительно менее активной дефосфорилированной форме (фосфорилаза b) (схема перехода изорм представлена на рисунке 2). Фосфорилаза а была получена в кристаллическом виде (мол. масса 190 кDa). Ее молекулы состоят из двух идентичных субъединиц, каждая из которых содержит существенный для каталитической активности остаток серина в фосфорилированной форме. Скорость превращения структурных единиц гликогена в глюкозо-1-фосфат регулируется в мышцах соотношением активной фосфорилазы а и менее активной фосфорилазы b . Взаимопревращения двух этих форм гликогенфосфорилазы происходят под действием специфичных ферментов, катализирующих процесс ковалентной модификации фосфорилазы. Фосфорилаза а превращается в менее активную фосфорилазу b под действием фермента, называемого фосфатазой фосфорилазы а; этот фермент, катализируя гидролитический разрыв связей, удаляет из молекулы фосфорилазы а фосфатные группы, необходимые для каталитической активности.
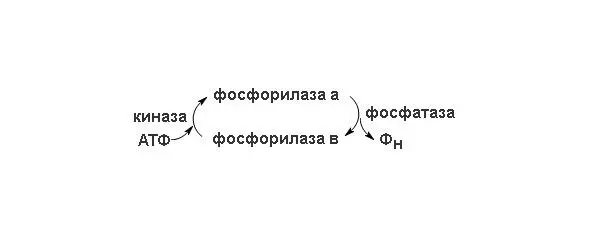
Рисунок 2. Схема перехода двух форм фосфорилазы.
Фосфорилаза b вновь превращается в активную фосфорилазу а под действием фермента, называемого киназой фосфорилазы b; он катализирует реакцию, в ходе которой АТФ фосфорилирует остатки серина в активном центре молекулы фосфорилазы b, что и приводит к образованию фосфорилазы а. Таким образом, благодаря действию двух ферментов, фосфатазы фосфорилазы а и киназы фосфорилазы b, соотношение активной фосфорилазы а и сравнительно мало активной фосфорилазы b в клетке может изменяться. В мышцах действует второй механизм регуляции гликогенфосфорилазной активности. Фосфорилаза b, сравнительно мало активная форма, может становиться более активной в результате нековалентного связывания с аллостерическим модулятором этого фермента, которым является AMФ; концентрация же AMФ в мышцах возрастает по мере распада АТФ в сократительных системах. Активации фосфорилазы b под действием AMФ препятствует АТФ, выступающий в роли отрицательного модулятора. Таким образом, активность фосфорилазы b определяется соотношением AMФ и ATФ. В отличие от фосфорилазы b фосфорилаза а не активируется AMФ; поэтому фосфорилазу а называют иногда AMФ-независимой формой, а фосфорилазу b AMФ-зависимой.
Читать дальшеИнтервал:
Закладка:
Похожие книги на «Биохимия метаболизма. Учебное пособие»
Представляем Вашему вниманию похожие книги на «Биохимия метаболизма. Учебное пособие» списком для выбора. Мы отобрали схожую по названию и смыслу литературу в надежде предоставить читателям больше вариантов отыскать новые, интересные, ещё непрочитанные произведения.












Обсуждение, отзывы о книге «Биохимия метаболизма. Учебное пособие» и просто собственные мнения читателей. Оставьте ваши комментарии, напишите, что Вы думаете о произведении, его смысле или главных героях. Укажите что конкретно понравилось, а что нет, и почему Вы так считаете.