Е. Бессолицына - Биохимия метаболизма. Учебное пособие
Здесь есть возможность читать онлайн «Е. Бессолицына - Биохимия метаболизма. Учебное пособие» — ознакомительный отрывок электронной книги совершенно бесплатно, а после прочтения отрывка купить полную версию. В некоторых случаях можно слушать аудио, скачать через торрент в формате fb2 и присутствует краткое содержание. ISBN: , Издательство: Литагент Ридеро, Жанр: Прочая научная литература, Биология, на русском языке. Описание произведения, (предисловие) а так же отзывы посетителей доступны на портале библиотеки ЛибКат.

- Название:Биохимия метаболизма. Учебное пособие
- Автор:
- Издательство:Литагент Ридеро
- Жанр:
- Год:неизвестен
- ISBN:9785448336638
- Рейтинг книги:3 / 5. Голосов: 1
-
Избранное:Добавить в избранное
- Отзывы:
-
Ваша оценка:
Биохимия метаболизма. Учебное пособие: краткое содержание, описание и аннотация
Предлагаем к чтению аннотацию, описание, краткое содержание или предисловие (зависит от того, что написал сам автор книги «Биохимия метаболизма. Учебное пособие»). Если вы не нашли необходимую информацию о книге — напишите в комментариях, мы постараемся отыскать её.
Биохимия метаболизма. Учебное пособие — читать онлайн ознакомительный отрывок
Ниже представлен текст книги, разбитый по страницам. Система сохранения места последней прочитанной страницы, позволяет с удобством читать онлайн бесплатно книгу «Биохимия метаболизма. Учебное пособие», без необходимости каждый раз заново искать на чём Вы остановились. Поставьте закладку, и сможете в любой момент перейти на страницу, на которой закончили чтение.
Интервал:
Закладка:
Гликолиз – процесс анаэробного окисления моносахаридов, в результате которого происходит синтез АТФ. У аэробных организмов АТФ синтезируется еще и в процессах клеточного дыхания, в которых утилизируются другие продукты гликолиза (пируват и NADH), поэтому направление реакций гликолиза сдвинуто в сторону образования продуктов метаболитического пути.
У анаэробных организмов гликолиз один из основных путей энергетического обмена углеводов, но утилизации остальных продуктов не происходит, накопление этих продуктов может привести к остановке гликолиза, а следовательно и остановке энергетического обмена (АТФ не образуется), поэтому необходимо утилизировать пируват и NADH. Для этого появилась «надстройка» – брожение, для окисления NADH и сдвига реакций в сторону образования пирувата.
Брожение
У аэробных организмов пируват и NADH, образовавшиеся в ходе гликолиза, утилизируются в ходе клеточного дыхания. У анаэробных организмов этого не происходит, поэтому необходима надстройка. Этой надстройкой является процесс брожения.
Брожение – это тип анаэробного окисления пирувата.
В настощее время описано множество путей брожения. Все они выявлены у прокариотических организмов. Но два типа бружения встречаются не только у прокариот, но и у эукариот. У эукариотических организмов обнаружены гомоферментативное молочнокислое и спиртовое брожение.
Гомоферментативное молочнокислое брожение
Гомоферментативное молочнокислое брожение получило свое название (схема представлена на рисунке 3), из-за того, что происходит одна реакция осуществляемая ферментом лактат дегидрогеназой молекула пирувата восстанавливается до лактата и донором протонов и электронов является NADH.
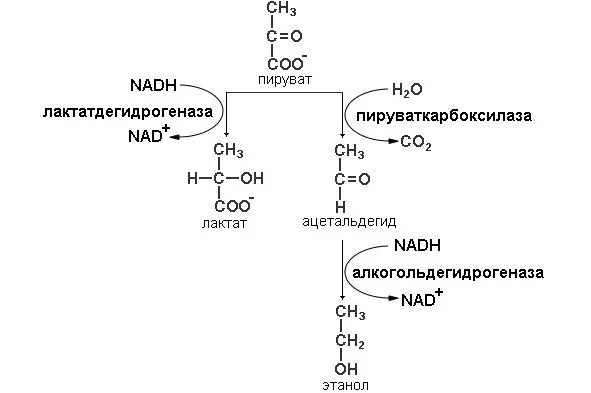
Рисунок 3: Схема реакций гомоферментативного молочнокислого и спиртового видов брожения
Данный тип брожения характерен для многих бактерий. Многие возбудители заболеваний, например, стафилококки и стрептококки, осуществляют этот тип брожения. Но данный брожения встречается и у полезных бактерий. Молочнокислые бактерии используются при изготовлении молочнокислых продуктов, сыров, сырокопченых колбас, при квашении капусты, засолке огурцов, силосовании при заготовке кормов. Также молочнокислое брожение встречается и у эукариот. Паразиты крови (простейшие, паразитирующие в крови млекопитающих (возбудители малярии, сонной болезни), также используют данный тип утилизации пирувата.
Лактатдегидрогеназа присутствует и в тканях млекопитающих. Хотя в обычных условиях наши мышцы получают вполне достаточные количества кислорода, чтобы произошло окисление пирувата и образование АТФ аэробным путем, бывают обстоятельства, когда поступление кислорода оказывается недостаточным. Например, при крайнем напряжении сил, когда уже весь запас кислорода израсходован, мышечные клетки образуют лактат путем брожения. Более того, в белых мышцах рыб или домашней птицы аэробный метаболизм относительно невелик, и основным конечным продуктом оказывается L-лактат. В организме человека есть такие ткани, которые слабо снабжаются кровью, например, хрусталик и роговица глаза. В клетках этих тканей окислительный метаболизм выражен слабо, а энергия в основном образуется при сбраживании глюкозы в лактат. Часть лактата, образующегося в мышцах и других тканях, поступает в кровь и переносится в печень, где он снова окисляется в пируват. Меньшая часть пирувата затем окисляется в цикле трикарбоновых кислот, но большая его часть снова превращается в глюкозу. Последняя может опять поступать в кровь и возвращаться в мышцы. Весь этот процесс называется циклом Кори.
Спиртовое брожение
У дрожжей и у других микроорганизмов, сбраживающих глюкозу не до лактата, а до этанола и СО 2, путь ферментативного расщепления глюкозы совпадает с описанным выше для анаэробного гликолиза на всем протяжении, за исключением этапа, катализируемого лактатдегидрогеназой (схема представлена на рисунке 3). В дрожжевых клетках, которые не содержат фермента, аналогичного лактатдегидрогеназе мышечной ткани, этот этап заменен двумя другими реакциями. В первой из них продукт расщепления глюкозы пируват теряет свою карбоксильную группу под действием пируватдекарбоксилазы. Эта реакция представляет собой простое декарбоксилирование; реального окисления пирувата при этом не происходит. Для проявления каталитической активности пируватдекарбоксилазе требуется Mg 2+. С молекулой этого фермента прочно связан кофермент тиаминпирофосфат. На последнем этапе спиртового брожения ацетальдегид восстанавливается до этанола за счет NADH, образовавшегося при окислении глицеральдегид-3-фосфата; эта реакция катализируется алкогольдегидрогеназой.
Читать дальшеИнтервал:
Закладка:
Похожие книги на «Биохимия метаболизма. Учебное пособие»
Представляем Вашему вниманию похожие книги на «Биохимия метаболизма. Учебное пособие» списком для выбора. Мы отобрали схожую по названию и смыслу литературу в надежде предоставить читателям больше вариантов отыскать новые, интересные, ещё непрочитанные произведения.












Обсуждение, отзывы о книге «Биохимия метаболизма. Учебное пособие» и просто собственные мнения читателей. Оставьте ваши комментарии, напишите, что Вы думаете о произведении, его смысле или главных героях. Укажите что конкретно понравилось, а что нет, и почему Вы так считаете.