Марк Мосевицкий - Распространненость жизни и уникальность разума?
Здесь есть возможность читать онлайн «Марк Мосевицкий - Распространненость жизни и уникальность разума?» — ознакомительный отрывок электронной книги совершенно бесплатно, а после прочтения отрывка купить полную версию. В некоторых случаях можно слушать аудио, скачать через торрент в формате fb2 и присутствует краткое содержание. Город: Санкт-Петербург, Год выпуска: 2008, ISBN: 2008, Издательство: ПИЯФ РАН, Жанр: Прочая научная литература, Физика, Биология, на русском языке. Описание произведения, (предисловие) а так же отзывы посетителей доступны на портале библиотеки ЛибКат.

- Название:Распространненость жизни и уникальность разума?
- Автор:
- Издательство:ПИЯФ РАН
- Жанр:
- Год:2008
- Город:Санкт-Петербург
- ISBN:5-86763-203-2
- Рейтинг книги:5 / 5. Голосов: 1
-
Избранное:Добавить в избранное
- Отзывы:
-
Ваша оценка:
Распространненость жизни и уникальность разума?: краткое содержание, описание и аннотация
Предлагаем к чтению аннотацию, описание, краткое содержание или предисловие (зависит от того, что написал сам автор книги «Распространненость жизни и уникальность разума?»). Если вы не нашли необходимую информацию о книге — напишите в комментариях, мы постараемся отыскать её.
Распространненость жизни и уникальность разума? — читать онлайн ознакомительный отрывок
Ниже представлен текст книги, разбитый по страницам. Система сохранения места последней прочитанной страницы, позволяет с удобством читать онлайн бесплатно книгу «Распространненость жизни и уникальность разума?», без необходимости каждый раз заново искать на чём Вы остановились. Поставьте закладку, и сможете в любой момент перейти на страницу, на которой закончили чтение.
Интервал:
Закладка:
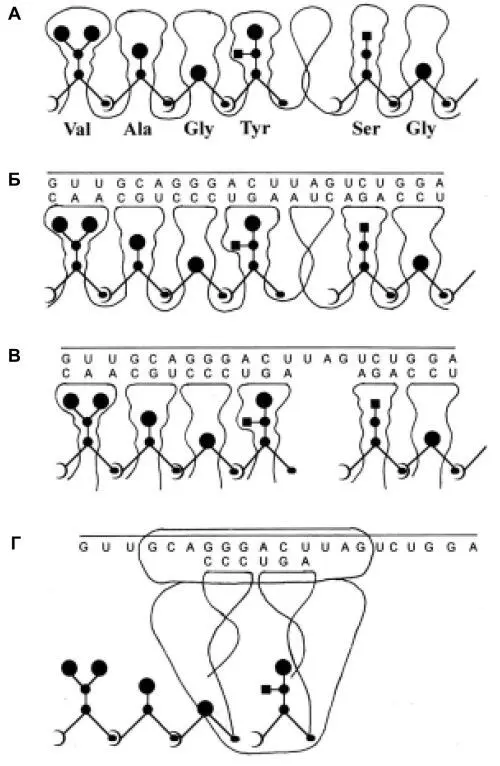
Рис. 1.
Рис. 1. Схемы, иллюстрирующие гипотезу перехода от примитивного синтеза предетерминированных пептидов к современному генетическому коду.
А. Образование белка (пептида), запрограммированного последовательностью петель-полостей единой молекулы РНК, связывающих активированные аминокислоты. Б. То же. Показана также “огибающая” РНК, нуклеотидная последовательность которой соответствует (комплементарна) ряду “торцов” петлевых элементов матрицы. В. Синтез пептидов на последовательности разделенных полостей-петель, набираемых из общего пула. Порядок петлевых элементов в наборе определяется связыванием их торцевых участков с соответствующими участками «огибающей» РНК, выполняющей таким образом роль информационной молекулы (мРНК). Соответственно, петлевые элементы в этой схеме являются предшественниками тРНК. Г. Эпизод современного синтеза белка на рибосоме: присоединение очередного звена к растущей белковой цепи. Вследствие изменения механизма узнавания адапторами соответствующих аминокислот петли тРНК перестали представлять собою «полости», хотя в них присутствуют необходимые для узнавания структурные элементы.
Схемы Рис. 1 предназначены для иллюстрации основных положений гипотезы и не отражают истинной конформации (укладки) матричных молекул РНК. Другие разъяснения приведены в тексте. Рисунок создан при участии В.В. Горбенко.
И это уже можно рассматривать как частичную реализацию информационных возможностей огибающих нитей. До полной реализации этих возможностей оставался один шаг: образование пула автореплицирующихся автономных РНК-адапторов, связывавших отдельные аминокислоты и узнававших соответствующие им участки в информационных нитях. Первоначально адапторные молекулы – предшественницы современных транспортных РНК (тРНК) – возникали как однопетлевые фрагменты РНК-матрицы. Сформировавшийся пул автономных РНК-петель обеспечивал связывание всех аминокислот, входивших тогда в состав пептидов. “Заряженные” аминокислотами автономные петли матричной РНК были также способны своими торцевыми участками подключаться к соответствующим (комплементарным) участкам огибающей РНК. Принципиально важным для дальнейшей эволюции качеством пула автономных адапторов аминокислот стала его универсальность: он мог быть использован при синтезе любого пептида в присутствии соответствующей огибающей РНК.
Переход к современному принципу кодирования аминокислотных последовательностей и к способу прочтения этого кода при синтезе белка в рибосоме иллюстрирует схема Рис. 1Б: связавшие аминокислоты петли-адапторы выстроены в том же порядке, что на Рис. 1A и Б, но не потому, что они в этом порядке связаны друг с другом в единой молекуле, а потому, что получили информацию о своей позиции благодаря взаимодействию с соответствующими участками огибающей РНК, принявшей на себя функции мРНК. Участки, узнаваемые в мРНК адапторными РНК, представляли собою кодоны, а их последовательность кодировала определенный пептид.
Поначалу новый способ кодирования белков развивался как вспомогательный вариант “под прикрытием” продолжавшего функционировать примитивного механизма. Главным эволюционным стимулом к переключению на новый способ кодирования белков явилась высокая рациональность этого способа, освободившего клетку от необходимости содержать и сохранять в поколениях значительное количество (по числу белков) РНК-матриц, размеры которых многократно превышали размеры соответствующих мРНК. После перехода к кодированию с участием мРНК открылась возможность увеличения как числа, так и размеров клеточных белков. Соответственно возросли их разнообразие и конформационная сложность, что позволило клеткам освоить новые пути метаболизма, усовершенствовать энергетику, кардинально повысить скорость и точность синтезов.
Первоначально характер связывания аминокислот соответствующими им адапторами сохранялся таким же, как при примитивном синтезе: полость, образованная петлей РНК, конформационно соответствовала аминокислоте и удерживала ее в связанном положении. Впоследствии механизм связывания был изменен. Оно стало осуществляться при посредничестве белковых ферментов аминоацил-тРНК-синтетаз, подключающих аминокислоты к 3'-концевому аденозину соответствующего адаптора (тРНК). При этом изменилось назначение петлевой области в молекулах тРНК. Ее структура сохранила специфичность, но преобразовалась соответственно новому назначению – служить объектом узнавания для специфической аминоацил-тРНК-синтетазы.
Читать дальшеИнтервал:
Закладка:
Похожие книги на «Распространненость жизни и уникальность разума?»
Представляем Вашему вниманию похожие книги на «Распространненость жизни и уникальность разума?» списком для выбора. Мы отобрали схожую по названию и смыслу литературу в надежде предоставить читателям больше вариантов отыскать новые, интересные, ещё непрочитанные произведения.










Обсуждение, отзывы о книге «Распространненость жизни и уникальность разума?» и просто собственные мнения читателей. Оставьте ваши комментарии, напишите, что Вы думаете о произведении, его смысле или главных героях. Укажите что конкретно понравилось, а что нет, и почему Вы так считаете.