Виктор Сбойчаков - Микробиология с основами эпидемиологии и методами микробиологических исследований
Здесь есть возможность читать онлайн «Виктор Сбойчаков - Микробиология с основами эпидемиологии и методами микробиологических исследований» — ознакомительный отрывок электронной книги совершенно бесплатно, а после прочтения отрывка купить полную версию. В некоторых случаях можно слушать аудио, скачать через торрент в формате fb2 и присутствует краткое содержание. ISBN: , Жанр: Медицина, Медицина, Прочая научная литература, Медицина, на русском языке. Описание произведения, (предисловие) а так же отзывы посетителей доступны на портале библиотеки ЛибКат.

- Название:Микробиология с основами эпидемиологии и методами микробиологических исследований
- Автор:
- Жанр:
- Год:неизвестен
- ISBN:978-5-299-00404-5
- Рейтинг книги:4 / 5. Голосов: 1
-
Избранное:Добавить в избранное
- Отзывы:
-
Ваша оценка:
Микробиология с основами эпидемиологии и методами микробиологических исследований: краткое содержание, описание и аннотация
Предлагаем к чтению аннотацию, описание, краткое содержание или предисловие (зависит от того, что написал сам автор книги «Микробиология с основами эпидемиологии и методами микробиологических исследований»). Если вы не нашли необходимую информацию о книге — напишите в комментариях, мы постараемся отыскать её.
Книга также будет полезна практическим лаборантам и фельдшерам, врачам микробиологам и студентам медицинских вузов.
Микробиология с основами эпидемиологии и методами микробиологических исследований — читать онлайн ознакомительный отрывок
Ниже представлен текст книги, разбитый по страницам. Система сохранения места последней прочитанной страницы, позволяет с удобством читать онлайн бесплатно книгу «Микробиология с основами эпидемиологии и методами микробиологических исследований», без необходимости каждый раз заново искать на чём Вы остановились. Поставьте закладку, и сможете в любой момент перейти на страницу, на которой закончили чтение.
Интервал:
Закладка:
Грибы размножаются как половым, так и бесполым (вегетативным) способом. Последний осуществляется с помощью простого митоза. Основные способы размножения:
1) фрагментация гифов мицелия грибов, в результате чего образуются артроспоры (рис. 15); если они образуют толстостенную оболочку, то называются хламидоспорами, высокоустойчивы к действию неблагоприятных факторов внешней среды;
2) почкование, в результате которого образуются бластоспоры;
3) образование бесполых спор, которое у одних грибов происходит в специальных вместилищах (спорангиях) — спорангиоспоры (эндоспоры); у других – на специализированных гифах-конидиеносцах — экзоспоры , или конидии (рис. 16).
Число, форма и размер конидий имеют особую организацию. Некоторые виды способны образовывать различные типы конидий – одноклеточные (микроконидии) либо многоклеточные (макроконидии). Бесполые структуры грибов называются анаморфами , а половые — телеоморфами. Последние наиболее значимы для систематики грибов. Половые стадии обнаружены у совершенных грибов, принадлежащих к классам Ascomycetes и Basidiomycetes , а также у немногочисленных представителей класса Zygomycetes . Процесс созревания половых спор у этих грибов происходит в специальных структурах: у аскоспор – в асках (сумках); у базидиоспор – на (в) плодовых телах.
У представителей класса Zygomycetes продуктом полового процесса, представляющего собой слияние коротких боковых ответвлений вегетативных гифов, являются одноклеточные образования – зигоспоры. Весь жизненный цикл несовершенных грибов ( Fungi imperfecti ) проходит в гаплоидной стадии. Они размножаются только бесполым путем.
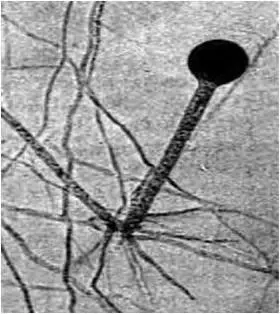
Рис. 16. Морфология грибов рода Rhizopus , класс Zygomycetes
Вирусы не способны к росту и бинарному делению, их размножение тесно связано с клеткой-хозяином. Процесс взаимодействия вирусов с чувствительной клеткой называется репродукцией . Выделяют раннюю и позднюю ее фазы. Ранняя фаза включает: 1) адсорбцию вириона на чувствительной клетке; 2) проникновение в клетку (пенетрацию); 3) раздевание вириона.
Начальные процессы адсорбции имеют неспецифический характер, в их основе может лежать электрическое взаимодействие положительно и отрицательно заряженных группировок на поверхности вируса и клетки. На адсорбцию влияют также рН, буферность и температура среды. При температуре 4 °C адсорбция носит синхронный характер, с повышением температуры скорость адсорбции увеличивается, но она приобретает асинхронный характер. Дальнейшее взаимодействие клеточных рецепторов и вирусных прикрепительных белков носит специфический характер.
Вирусы используют рецепторы, предназначенные для проникновения в клетку необходимых для ее жизнедеятельности веществ: гормонов, ферментов, факторов роста, других питательных веществ. Клеточные рецепторы имеют разную химическую природу. Так, для вируса гриппа и парагриппа рецепторами являются структуры, содержащие сиаловую (нейраминовую) кислоту.
Прикрепление вириона к клеточной поверхности осуществляется следующим образом. Вначале происходит образование единичной связи прикрепительного белка с рецептором – обратимая адсорбция. В этот момент, изменяя рН среды, воздействуя ультразвуком, антителами, можно удалить вирион с поверхности клетки. Для того чтобы наступила необратимая адсорбция, должны появиться множественные связи между вирионами и клеточными рецепторами. Число молекул клеточных рецепторов, участвующих в адсорбции, может доходить до 3000.
Прикрепительные белки вирусов могут находиться в составе уникальных образований, таких, как фибры у аденовирусов. У сложно организованных вирусов эти белки входят в состав шипов на поверхности суперкапсиды, например, у вируса гриппа имеется 300 – 450 шипов гемагглютинина. Просто организованные вирусы содержат прикрепительные белки в составе капсида.
Проникновение (пенетрация) вирусов в клетку осуществляется за счет двух механизмов, взаимодополняющих друг друга: виропексиса (эндоцитоза) и слияния вирусной и клеточной мембраны.
Термин «виропексис», предложенный в 1948 г. Фазекасом де Сан Гро, означает, что вирион попадает в цитоплазму в результате инвагинации участка плазматической мембраны и образования вакуоли. Виропексис является частным случаем рецепторного эндоцитоза, который обеспечивает поступление в клетку аминокислот, нуклеотидов, гормонов и других веществ из межклеточной жидкости. Большинство вирусов проникает в клетку путем рецепторного эндоцитоза, некоторые вирусы – за счет механизма слияния. Функцию белка слияния у вируса гриппа выполняет малая гемагглютинирующая субъединица HA2, у вирусов парагриппа белок слияния — fusion protein (англ. fusiоn – слияние).
Читать дальшеИнтервал:
Закладка:
Похожие книги на «Микробиология с основами эпидемиологии и методами микробиологических исследований»
Представляем Вашему вниманию похожие книги на «Микробиология с основами эпидемиологии и методами микробиологических исследований» списком для выбора. Мы отобрали схожую по названию и смыслу литературу в надежде предоставить читателям больше вариантов отыскать новые, интересные, ещё непрочитанные произведения.












Обсуждение, отзывы о книге «Микробиология с основами эпидемиологии и методами микробиологических исследований» и просто собственные мнения читателей. Оставьте ваши комментарии, напишите, что Вы думаете о произведении, его смысле или главных героях. Укажите что конкретно понравилось, а что нет, и почему Вы так считаете.