Виктор Сбойчаков - Микробиология с основами эпидемиологии и методами микробиологических исследований
Здесь есть возможность читать онлайн «Виктор Сбойчаков - Микробиология с основами эпидемиологии и методами микробиологических исследований» — ознакомительный отрывок электронной книги совершенно бесплатно, а после прочтения отрывка купить полную версию. В некоторых случаях можно слушать аудио, скачать через торрент в формате fb2 и присутствует краткое содержание. ISBN: , Жанр: Медицина, Медицина, Прочая научная литература, Медицина, на русском языке. Описание произведения, (предисловие) а так же отзывы посетителей доступны на портале библиотеки ЛибКат.

- Название:Микробиология с основами эпидемиологии и методами микробиологических исследований
- Автор:
- Жанр:
- Год:неизвестен
- ISBN:978-5-299-00404-5
- Рейтинг книги:4 / 5. Голосов: 1
-
Избранное:Добавить в избранное
- Отзывы:
-
Ваша оценка:
Микробиология с основами эпидемиологии и методами микробиологических исследований: краткое содержание, описание и аннотация
Предлагаем к чтению аннотацию, описание, краткое содержание или предисловие (зависит от того, что написал сам автор книги «Микробиология с основами эпидемиологии и методами микробиологических исследований»). Если вы не нашли необходимую информацию о книге — напишите в комментариях, мы постараемся отыскать её.
Книга также будет полезна практическим лаборантам и фельдшерам, врачам микробиологам и студентам медицинских вузов.
Микробиология с основами эпидемиологии и методами микробиологических исследований — читать онлайн ознакомительный отрывок
Ниже представлен текст книги, разбитый по страницам. Система сохранения места последней прочитанной страницы, позволяет с удобством читать онлайн бесплатно книгу «Микробиология с основами эпидемиологии и методами микробиологических исследований», без необходимости каждый раз заново искать на чём Вы остановились. Поставьте закладку, и сможете в любой момент перейти на страницу, на которой закончили чтение.
Интервал:
Закладка:
Культура бактерий в ограниченном объеме питательной среды называется периодической, и описанные фазы роста характерны именно для таких культур. Если благоприятные для роста бактерий условия поддерживать путем подачи свежей питательной среды и оттока продуктов распада, то их культивирование станет непрерывным. Это происходит при различных биотехнологических процессах.
По типу дыхания риккетсии являются аэробами. Их характерными метаболическими признаками являются: полное окисление глутаминовой кислоты и отсутствие утилизации глюкозы. Риккетсии имеют белковые эндотоксины, чувствительные к формалину.
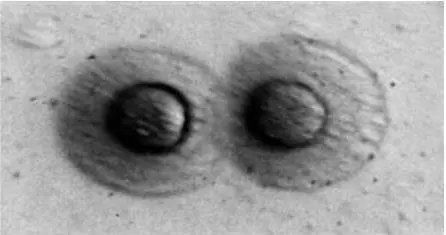
Рис. 13. Колонии микоплазм
Риккетсии размножаются путем обычного деления кокковидных и палочковидных форм с последующей фазой гомогенных популяций. Другой формой размножения является мицеллярное (как у грибов) дробление нитевидных форм. Нитевидные формы соответствуют наиболее ранней фазе развития риккетсий, а кокковидные и короткие палочковидные формы представляют конечную стадию их деления. Риккетсии размножаются гораздо медленнее, чем бактерии.
Размножение микоплазм не может происходить без участия стерола. Хотя микоплазмы и могут расти на искусственных питательных средах, для своего роста они нуждаются также в холестерине, жирных кислотах и нативном белке. Микоплазмы проникают через водную пленку и адсорбируются агаром. Через 18 ч внутри агара формируются маленькие сферические колонии, через 48 ч эта колония достигает поверхности водной пленки агара. Внешние колонии микоплазм имеют вид яичницы-глазуньи (рис. 13).
Хламидии существуют в двух формах, различающихся по морфологическим и биологическим свойствам. Инфекционной внеклеточной формой является элементарное тельце (ЭТ), а вегетативной, репродуцирующейся, внутриклеточной – ретикулярное тельце (РТ). Элементарное тельце имеет вид сферы диаметром 0,15 – 0,2 мкм, ретикулярное – имеет структуру типичных грамотрицательных бактерий размером около 1 мкм (рис. 14). В ЭТ содержится больше дисульфидных связей, что позволяет им противостоять осмотическому давлению.
Первый этап инфекционного процесса – адсорбция ЭТ на плазмалемме клетки-хозяина. Важную роль здесь играют электростатические силы. Внедрение хламидий происходит путем эндоцитоза. Инвагинация участка плазмалеммы с адсорбированным ЭТ происходит в цитоплазму с образованием фагоцитарной вакуоли. Эта фаза занимает 7 – 10 ч, затем в течение 6 – 8 ч происходит реорганизация ЭТ в вегетативную форму – ретикулярное тельце, способное к росту и делению. Именно в этой фазе эффективно применение антибактериальных препаратов, поскольку ЭТ к ним не чувствительно.
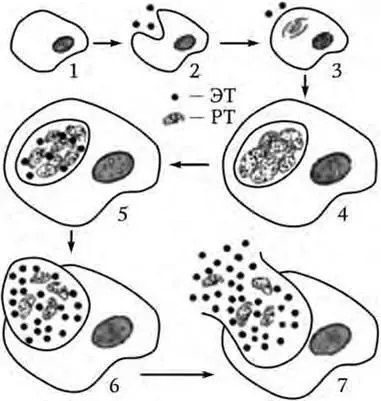
Рис. 14. Цикл размножения хламидий (стадии развития от попадания ЭТ в клетку до выхода ЭТ следующего поколения):
1 – адсорбция элементарного тельца; 2 – проникновение элементарного тельца в клетку; 3 – реорганизация элементарного тельца в ретикулярное тельце; 4 – деление ретикулярного тельца; 5 – созревание ретикулярных телец в элементарные; 6 – накопление ретикулярных телец в эндосоме; 7 – выход хламидий из клетки
Размножение хламидий приводит к формированию включений, известных под названием телец Провачека. В течение 18 – 24 ч развития они локализованы в цитоплазматическом пузырьке, образованном из мембраны клетки-хозяина. Во включении может содержаться от 100 до 500 хламидий. Остановка процесса на этой стадии ведет к персистенции хламидийной инфекции. Далее начинается процесс созревания ретикулярных телец через переходные (промежуточные) тельца в течение 36 – 42 ч развития в ЭТ следующего поколения. Полный цикл репродукции хламидий равен 48 – 72 ч и завершается разрушением пораженной клетки. В случае возникновения для хламидий неблагоприятных метаболических условий этот процесс может затягиваться на более длительный период.
Хламидии могут высвобождаться из инфицированной клетки через узкий ободок цитоплазмы. При этом клетка может сохранять жизнеспособность, этим можно объяснить бессимптомность течения хламидийной инфекции.
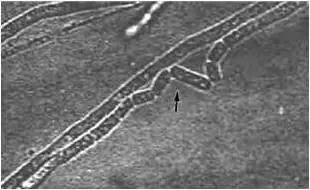
Рис. 15. Артроспоры культуры гриба Trichosporon beigelii
Читать дальшеИнтервал:
Закладка:
Похожие книги на «Микробиология с основами эпидемиологии и методами микробиологических исследований»
Представляем Вашему вниманию похожие книги на «Микробиология с основами эпидемиологии и методами микробиологических исследований» списком для выбора. Мы отобрали схожую по названию и смыслу литературу в надежде предоставить читателям больше вариантов отыскать новые, интересные, ещё непрочитанные произведения.












Обсуждение, отзывы о книге «Микробиология с основами эпидемиологии и методами микробиологических исследований» и просто собственные мнения читателей. Оставьте ваши комментарии, напишите, что Вы думаете о произведении, его смысле или главных героях. Укажите что конкретно понравилось, а что нет, и почему Вы так считаете.