Николай Воронцов - Развитие эволюционных идей в биологии
Здесь есть возможность читать онлайн «Николай Воронцов - Развитие эволюционных идей в биологии» — ознакомительный отрывок электронной книги совершенно бесплатно, а после прочтения отрывка купить полную версию. В некоторых случаях можно слушать аудио, скачать через торрент в формате fb2 и присутствует краткое содержание. Город: Москва, Год выпуска: 1999, ISBN: 1999, Издательство: Прогресс-Традиция, Жанр: История, Биология, на русском языке. Описание произведения, (предисловие) а так же отзывы посетителей доступны на портале библиотеки ЛибКат.

- Название:Развитие эволюционных идей в биологии
- Автор:
- Издательство:Прогресс-Традиция
- Жанр:
- Год:1999
- Город:Москва
- ISBN:5-88800-107-4
- Рейтинг книги:4 / 5. Голосов: 1
-
Избранное:Добавить в избранное
- Отзывы:
-
Ваша оценка:
Развитие эволюционных идей в биологии: краткое содержание, описание и аннотация
Предлагаем к чтению аннотацию, описание, краткое содержание или предисловие (зависит от того, что написал сам автор книги «Развитие эволюционных идей в биологии»). Если вы не нашли необходимую информацию о книге — напишите в комментариях, мы постараемся отыскать её.
В книге подробно прослежено развитие эволюционной идеи, возникшей за тысячи лет до Дарвина и принадлежащей к числу немногих общенаучных фундаментальных идей, определивших мышление юнца XIX и XX столетия. Проанализированы все этапы зарождения и формирования представлений об эволюции, начиная с первобытного общества. Особое внимание уделено истокам, развитию и восприятию дарвинизма, в частности, в России, влиянию дарвинизма на все естествознание.
Последние главы показывают, как сегодняшние открытия в области молекулярной биологии, генетики и многих других дисциплин готовят почву для нового синтеза в истории эволюционизма.
Книга насыщена массой интересных и поучительных исторических подробностей, как правило, малоизвестных, и содержит большое число иллюстраций, как авторских, так и взятых из труднодоступных изданий. Книга рассчитана на широкого читателя, не только биолога, но любого, интересующегося современной наукой ее историей.
Развитие эволюционных идей в биологии — читать онлайн ознакомительный отрывок
Ниже представлен текст книги, разбитый по страницам. Система сохранения места последней прочитанной страницы, позволяет с удобством читать онлайн бесплатно книгу «Развитие эволюционных идей в биологии», без необходимости каждый раз заново искать на чём Вы остановились. Поставьте закладку, и сможете в любой момент перейти на страницу, на которой закончили чтение.
Интервал:
Закладка:
При дивергенции видов по частным признакам путем точковых мутаций общность генного состава разных видов, родов, семейств и даже отрядов может сохраняться достаточно долго. Завоевание новой адаптивной зоны обычно сопряжено с приобретением принципиально новых признаков, неким «подъемом организации», связанным с коренной перестройкой генома и, соответственно, всей биохимии, физиологии и морфологии организмов. В этом случае в гомологичных участках геномов разных видов могут происходить гомологичные мутации, которые легко подхватываются сходно направленным естественным отбором; в результате переход в другую адаптивную зону (например, завоевание суши амфибиями, окончательный отрыв от водной среды у рептилий или приобретение теплокровности, четырехкамерного сердца, новой структуры черепа у предков млекопитающих) может затрагивать одновременно не один, а многие виды, принадлежащие даже к разным семействам и отрядам исходного класса [576] Воронцов Н. Н, О гомологической изменчивости // Проблемы кибернетики. Т. 16. М., 1966, с. 221-229; Воронцов Н. Н. Эволюция пищеварительной системы грызунов. Новосибирск: Наука, 1967, 240 с.
.
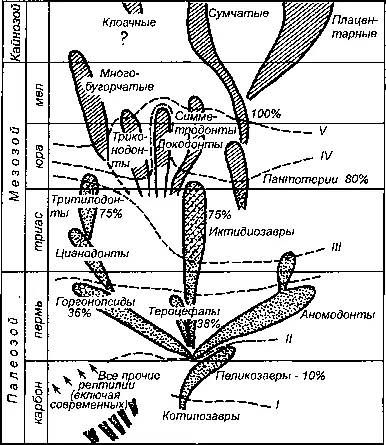
Рис. 253. Параллельное возникновение признаков млекопитающих в разных группах зверообразных рептилий и у мезозойских млекопитающих. Проценты показывают долю характерных для млекопитающих признаков в разных ископаемых группах рептилий. Римскими цифрами обозначены эволюционные уровни продвижения (грады) от рептилий до млекопитающих. Ясно, что млекопитающие относительно просто очерчиваются как уровень (града), но не могут быть очерчены как ветвь (клада). Филогения по Симпсону, процент сходства по Кун-Шнидеру.
Из Н. И. Воронцова (1980).
Многочисленные примеры парафилии на ботаническом материале приводит Н. Н. Цвелев. По мнению этого компетентного специалиста, как современные голосеменные, так и покрытосеменные возникли парафилетически [577] Цвелев Н. Н. О происхождении и основных направлениях эволюции злаков (Роасеае) // Проблемы эволюции, т. 4. Новосибирск: Наука, 1975, с. 107-117; Он же. О значении дивергенции и конвергенции в эволюции организмов // Вопросы развития эволюционной теории в XX веке. Л.: Наука, 1979, с. 23-31.
.
В случае подобного парафилетического происхождения таксонов высокого ранга в результате параллельного преобразования геномов разных видов мы не можем отрицать того, что в основе парафилии все же лежит дивергенция, ибо сама возможность гомологичного мутирования и параллельного развития вызвана в конечном счете общностью их происхождения.
Роль вирусной трансдукции в макроэволюции. Проблема горизонтального переноса
Еще в 1957 г. К. Уодингтон оценил эволюционное значение открытия трансдукции (переноса) бактериальных генов от одного вида бактерий к другому через инфицирование бактериофагом и уже тогда предположил возможность существования подобного механизма переноса генетической информации не только у прокариот, к которым относятся бактерии, но и у эукариот. Следует признать, что эволюционисты до недавнего времени предпочитали не знать или не пытались оценить революционизирующее значение этих фактов для эволюционной теории. Как справедливо отметил С. В. Мейен в выступлении в дискуссии по докладу автора в Геологическом институте АН СССР, для теории не важно, имеем ли мы дело с редким или частым явлением, для теории важно, существует ли вообще данное явление или нет.
Те самые методы преобразования генома эукариот, которые ныне применяются в опытах по генной инженерии, могли быть использованы и природой в течение 3,5 млрд. лет органической эволюции. Вопрос о том, сколь часто могла сохраниться клетка организма, в которую с помощью вирусной трансдукции был внесен участок генома другого вида, рода или отряда, — вопрос количественный, а не качественный. Сколь бы редко не происходил этот процесс, для нас важно, что он принципиально мог происходить, что новый геном мог синтезироваться не только путем симбиогенеза, но и путем трансдукции.
Наиболее увлекательный пример горизонтального переноса между представителями разных подимперий органического мира — прокариотами и эукариотами — был открыт в 1981 г. Дж. Мартином и И. Фридовичем. Есть семейство сребробрюшковых рыб (Leiogna thidae ) со светящимся брюшком. Его свечение обеспечивает симбиотическая биолюминесцирующая бактерия Photobacter leiognathi. Этот вид наряду с типичным для прокариот ферментом-металлопротеидом — Fe-супероксидисмутазой — имеет, как и рыбы, типичный для эукариот металлопротеид — Cu-Zn-супероксидисмутазу. Сравнение аминокислотного состава этого белка у данного вида фотобактерий и у сребробрюшковых рыб показало их сходство друг с другом, но не с другими металлопротеидами прокариот. Естественно предположение о горизонтальном переносе данного гена от рыб к симбиотическим бактериям!
Читать дальшеИнтервал:
Закладка:
Похожие книги на «Развитие эволюционных идей в биологии»
Представляем Вашему вниманию похожие книги на «Развитие эволюционных идей в биологии» списком для выбора. Мы отобрали схожую по названию и смыслу литературу в надежде предоставить читателям больше вариантов отыскать новые, интересные, ещё непрочитанные произведения.












Обсуждение, отзывы о книге «Развитие эволюционных идей в биологии» и просто собственные мнения читателей. Оставьте ваши комментарии, напишите, что Вы думаете о произведении, его смысле или главных героях. Укажите что конкретно понравилось, а что нет, и почему Вы так считаете.