Николай Воронцов - Развитие эволюционных идей в биологии
Здесь есть возможность читать онлайн «Николай Воронцов - Развитие эволюционных идей в биологии» — ознакомительный отрывок электронной книги совершенно бесплатно, а после прочтения отрывка купить полную версию. В некоторых случаях можно слушать аудио, скачать через торрент в формате fb2 и присутствует краткое содержание. Город: Москва, Год выпуска: 1999, ISBN: 1999, Издательство: Прогресс-Традиция, Жанр: История, Биология, на русском языке. Описание произведения, (предисловие) а так же отзывы посетителей доступны на портале библиотеки ЛибКат.

- Название:Развитие эволюционных идей в биологии
- Автор:
- Издательство:Прогресс-Традиция
- Жанр:
- Год:1999
- Город:Москва
- ISBN:5-88800-107-4
- Рейтинг книги:4 / 5. Голосов: 1
-
Избранное:Добавить в избранное
- Отзывы:
-
Ваша оценка:
Развитие эволюционных идей в биологии: краткое содержание, описание и аннотация
Предлагаем к чтению аннотацию, описание, краткое содержание или предисловие (зависит от того, что написал сам автор книги «Развитие эволюционных идей в биологии»). Если вы не нашли необходимую информацию о книге — напишите в комментариях, мы постараемся отыскать её.
В книге подробно прослежено развитие эволюционной идеи, возникшей за тысячи лет до Дарвина и принадлежащей к числу немногих общенаучных фундаментальных идей, определивших мышление юнца XIX и XX столетия. Проанализированы все этапы зарождения и формирования представлений об эволюции, начиная с первобытного общества. Особое внимание уделено истокам, развитию и восприятию дарвинизма, в частности, в России, влиянию дарвинизма на все естествознание.
Последние главы показывают, как сегодняшние открытия в области молекулярной биологии, генетики и многих других дисциплин готовят почву для нового синтеза в истории эволюционизма.
Книга насыщена массой интересных и поучительных исторических подробностей, как правило, малоизвестных, и содержит большое число иллюстраций, как авторских, так и взятых из труднодоступных изданий. Книга рассчитана на широкого читателя, не только биолога, но любого, интересующегося современной наукой ее историей.
Развитие эволюционных идей в биологии — читать онлайн ознакомительный отрывок
Ниже представлен текст книги, разбитый по страницам. Система сохранения места последней прочитанной страницы, позволяет с удобством читать онлайн бесплатно книгу «Развитие эволюционных идей в биологии», без необходимости каждый раз заново искать на чём Вы остановились. Поставьте закладку, и сможете в любой момент перейти на страницу, на которой закончили чтение.
Интервал:
Закладка:
В чем состоит принципиальное отличие симгенеза (синтезогенеза) от сегрегациогенеза? При анагенезе и кладогенезе процесс видообразования может носить (но не обязательно) постепенный, градуалисгический характер. В этом случае можно говорить о том, что налэпление отдельных мутаций структурных генов, обеспечиваемое как отбором, так и генетико-автоматическими процессами (а нередко и совместным действием этих двух формирующих факторов эволюции), может идти постепенно. Так же постепенно (нередко вообще как побочный эффект дивергенции по другим признакам) формируются барьеры изолирующих механизмов эволюции, которые со временем могут стать абсолютными. Таким образом, в случае анагенеза и кладогенеза макроэволюция действительно может идти через микроэволюцию в соответствии с одним из постулатов СТЭ.
Иначе обстоит дело при симгенезе. Даже гибридогенное видообразование, не говоря уже о случаях образования таксонов более высокого ранга путем симгенеза, не может быть сведено к классическим микроэволюционным процессам, здесь нет элементарного эволюционного явления — изменения генотипического состава популяции, ибо гибридная популяция ранее репродуктивно изолированных видов есть нечто принципиально новое: если в случае анагенеза и кладогенеза отбор, изоляция и генетико-автоматические процессы вызывают и направляют эволюцию, то в случае синтезогенеза инициирующим фактором является объединение разных геномов.
Бесспорно, что это объединение разных геномов и создание нового сбалансированного генома идет на фоне действия естественного отбора, отбрасывающего нежизнеспособные комбинации геномов. Но как элиминируемые, так и отбираемые индивидуумы в случае гибридогенеза оказываются изначально репродуктивно изолированными от родительских видов. Таким образом, синтезогенез должен рассматриваться (в случае возникновения таксона ранга вида и выше за счет объединения геномов разных видов) как макроэволюционный процесс, не вызванный микроэволюционными процессами. Вместе с тем ясно, что вслед за возникновением макроэволюционного акта начнут действовать на уровне новой генетической системы микро-эволюционные процессы.
Наиболее понятна и абсолютно доказана роль симгенеза на низших этапах макроэволюции, на уровне образования таксонов ранга вида, рода. Вместе с тем накапливаются данные, говорящие о том, что и мегаэволюция может идти за счет процессов интеграции разнородных геномов. Мы рассмотрим эти два уровня интеграции геномов порознь.
Симгенез в макроэволюции (видообразование)
Экспериментальный синтез видов
Возможность гибридогенного происхождения отдельных видов допускалась еще Линнеем и другими ботаниками того времени. Однако экспериментальные доказательства такой возможности и пути преодоления стерильности межвидовых гибридов были открыты лишь в 20—30-х гг. XX века.
Общеизвестны классические опыты Георгия Дмитриевича Карпеченко (1899—1942) по синтезу межродового гибрида между редькой (Raphanus sativus ) и капустой (Brassica oleracea ). При гибридизации (рис. 239) геномы редьки RR и капусты ВВ, имеющие сходное число хромосом (2n=18), дают гаплоидные гаметы с 9 хромосомами типа Л и В; их слияние дает стерильный гибрид с геномом RB. Стерильность вызвана нарушением конъюгации хромосом в мейозе, поскольку хромосомы группы R не имеют гомологов в группе В. В результате нарушений мейоза образовывались единичные гаметы с нередуцированным числом хромосом с геномом RB. Слияние двух таких гамет ведет к возникновению аллополиплоида с геномом RRBB.
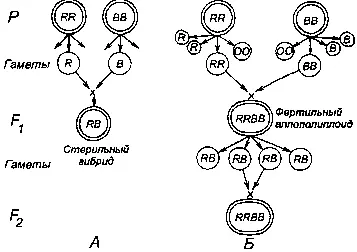
Рис. 239. Схема экспериментов Г. Д. Карпеченко по синтезу рафанобрассики — межродового гибрида с неограниченной плодовитостью, репродуктивно изолированного от предковых родов — редьки (Raphanus ) и капусты (Brassica ).
Из: Н. Н. Воронцов (1980).
У полученного аллополиплоида в мейозе хромосомы R конъюгировали с if, а В с В. Таким образом, была впервые в мире преодолена гибридная стерильность. Эта работа Г. Д. Карпеченко, результаты которой были опубликованы в 1924—1927 гг., не только показала способ получения плодовитых гибридов при отдаленной гибридизации, что затем стало широко использоваться в селекционной практике всего мира, но и продемонстрировала пути видообразования за счет аллополиплоидии в мире растений. Не случайно итоговая работа 1927 г. имела подзаголовок «К проблеме экспериментального видообразования» [500] Карпеченко Г. Д. Избранные труды. М.: Наука 1971, 304 с.
. По сути дела, синтез рафанобрассики был первым случаем конструирования нового генома, т. е. того, что в конце 70-х гг. нашего века стало называться генетической инженерией.
Интервал:
Закладка:
Похожие книги на «Развитие эволюционных идей в биологии»
Представляем Вашему вниманию похожие книги на «Развитие эволюционных идей в биологии» списком для выбора. Мы отобрали схожую по названию и смыслу литературу в надежде предоставить читателям больше вариантов отыскать новые, интересные, ещё непрочитанные произведения.












Обсуждение, отзывы о книге «Развитие эволюционных идей в биологии» и просто собственные мнения читателей. Оставьте ваши комментарии, напишите, что Вы думаете о произведении, его смысле или главных героях. Укажите что конкретно понравилось, а что нет, и почему Вы так считаете.