Николай Воронцов - Развитие эволюционных идей в биологии
Здесь есть возможность читать онлайн «Николай Воронцов - Развитие эволюционных идей в биологии» — ознакомительный отрывок электронной книги совершенно бесплатно, а после прочтения отрывка купить полную версию. В некоторых случаях можно слушать аудио, скачать через торрент в формате fb2 и присутствует краткое содержание. Город: Москва, Год выпуска: 1999, ISBN: 1999, Издательство: Прогресс-Традиция, Жанр: История, Биология, на русском языке. Описание произведения, (предисловие) а так же отзывы посетителей доступны на портале библиотеки ЛибКат.

- Название:Развитие эволюционных идей в биологии
- Автор:
- Издательство:Прогресс-Традиция
- Жанр:
- Год:1999
- Город:Москва
- ISBN:5-88800-107-4
- Рейтинг книги:4 / 5. Голосов: 1
-
Избранное:Добавить в избранное
- Отзывы:
-
Ваша оценка:
Развитие эволюционных идей в биологии: краткое содержание, описание и аннотация
Предлагаем к чтению аннотацию, описание, краткое содержание или предисловие (зависит от того, что написал сам автор книги «Развитие эволюционных идей в биологии»). Если вы не нашли необходимую информацию о книге — напишите в комментариях, мы постараемся отыскать её.
В книге подробно прослежено развитие эволюционной идеи, возникшей за тысячи лет до Дарвина и принадлежащей к числу немногих общенаучных фундаментальных идей, определивших мышление юнца XIX и XX столетия. Проанализированы все этапы зарождения и формирования представлений об эволюции, начиная с первобытного общества. Особое внимание уделено истокам, развитию и восприятию дарвинизма, в частности, в России, влиянию дарвинизма на все естествознание.
Последние главы показывают, как сегодняшние открытия в области молекулярной биологии, генетики и многих других дисциплин готовят почву для нового синтеза в истории эволюционизма.
Книга насыщена массой интересных и поучительных исторических подробностей, как правило, малоизвестных, и содержит большое число иллюстраций, как авторских, так и взятых из труднодоступных изданий. Книга рассчитана на широкого читателя, не только биолога, но любого, интересующегося современной наукой ее историей.
Развитие эволюционных идей в биологии — читать онлайн ознакомительный отрывок
Ниже представлен текст книги, разбитый по страницам. Система сохранения места последней прочитанной страницы, позволяет с удобством читать онлайн бесплатно книгу «Развитие эволюционных идей в биологии», без необходимости каждый раз заново искать на чём Вы остановились. Поставьте закладку, и сможете в любой момент перейти на страницу, на которой закончили чтение.
Интервал:
Закладка:
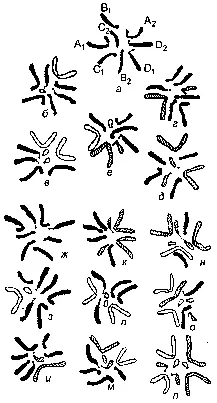
Рис. 203. Схема получения кариотипа нового синтезированного вида Crepis nova :
а — хромосомы Cr. tectorum; б, г — исходные гетерозиготные транслоканты; в, д — те же транслокацни, но переведенные в гомозиготное состояние; г — гибрид между в и д; ж-п — девять возможных комбинаций хромосом в потомстве е; п — Cr. nova.
Е. Н. Герасимовой из: Н. Н. Воронцов.
Идеи Четверикова роли явления рецессивности в эволюции с помощью Н. В. Тимофеева-Ресовского и Ф. Г. Добржанского распространились по мировому сообществу генетиков и эволюционистов и стали восприниматься как самоочевидные. В нашей стране на эти идеи особое внимание обратил Иван Иванович Шмальгаузен (1884—1963). В самом деле, ведь отбор идет по фенотипам, и, таким образом, особи с генотипами ААи Аа, имея одинаковый фенотип благодаря доминантности аллеля Анад рецессивным аллелем а, обладают равной селективной ценностью перед лицом естественного отбора. Таким образом, под покровом доминантного аллеля Арецессивный аллель а может очень долго существовать в популяции даже в том случае, если гомозиготы по рецессивному аллелю ааимеют пониженную жизнеспособность (табл. 3).
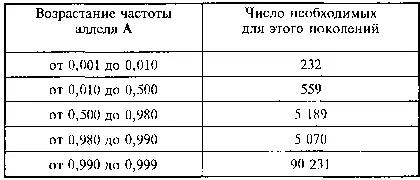
Как видно из теоретических расчетов, полученных в 1939 г. немецким теоретиком К. Пэтау [382] PStau К. Zeitschr fur induktive Abstammungs- und Vererbungslehre, 1939, B. 76, S. 220-238.
, нарастание частоты доминантного аллеля в случае его селективной ценности вначале идет относительно быстро, но затем замедляется и не приводит на протяжении многих тысяч поколений к полному исчезновению селективно не выгодного рецессивного аллеля. При смене условий, в случае, если генотип аа будет иметь повышенную селективную ценность в сравнении с генотипами АА и Аа, повышение частоты аллеля а пойдет в обратном направлении в сравнении с рассмотренным в таблице и более быстрыми темпами. Это явление существования в популяции рецессивных аллелей в гетерозиготном состоянии было названо И. И. Шмальгаузеном и С. М. Гершензоном [383] Г зршензон C. M. «Мобилизационный резерв» внутривидовой изменчивости // Журн. общ. биол., 1941, т. 2, №1.; Шмальгаузен И. И. Факторы эволюции. К теории стабилизирующего отбора. М.: Изд-во АН СССР, 1946.
в терминологии межвоенной поры «скрытым мобилизационным резервом внутривидовой изменчивости».
Генетикам-популяционистам того времени все более убедительным представлялось сведение микроэволюционного процесса к процессу постепенной смены генных частот, точнее, частот аллелей. Но поскольку и макроэволюция представлялась в ту пору бесспорным продолжением и развитием микроэволюции, то и вся эволюция стала постепенно восприниматься как процесс неуклонного и постепенного изменения частот аллелей. В результате к моменту возникновения синтетической теории эволюции генетикам-популяционистам стал представляться бесспорным процесс постепенности смены частот, т. е. постепенность стала казаться неотделимой чертой эволюционного процесса. Таким образом, произошел на новом уровне возврат к лейбницевскому «закону непрерывности», к дарвиновскому градуализму. Интересно, что на этот раз возврат к градуализму произошел в среде генетиков, среди которых еще за 20—30 лет до этого господствовали макромутационистские представления.
Однако градуализм преобладал лишь среди генетиков-популяционистов, притом связанных с изучением животных объектов. В 20—30-е годы стало ясно, что в видообразовании растений особую роль играет кратное увеличение числа хромосом — полиплоидия. В объединении идей эволюционной цитогенетики и ботаники участвовали исследователи многих стран. В СССР это были, в первую очередь, ученые школы Николая Ивановича Вавилова (рис. 204): Г. А. Левитский (рис. 205), от которого идут термины «кариотип» и «идеограмма» [384] Левитский Г. А. Материальные основы наследственности. Киев: Госиздат УкрСССР, 1924; Левитский Г. А. Карио- и генотипические изменения в процессе эволюции // Тр. по прикладной ботанике, генетике и селекции, 1925, т. 16, вып. 5; Левитский Г. А. Цитологические основы эволюции // Природа, 1939, №5. Переиздания см.: Левитский Г. А. Цитология растений. Избр. труды. М.: Наука, 1976, 351 с.
; Г. Д. Карпеченко (рис. 206), впервые получивший плодовитый межродовой гибрид — рафанобрассику [385] Карпеченко Г. Д. Полиплоидные гибриды Raphanus satrivus L. х Brassica oleracea L. // Tp. по прикл. ботанике, генет. и селекции, 1927, т. 17, вып. 3, с. 305-410; Polyploid hybrids of Raphanus satrivus L. x Brassica oleracea L. // Zeitschr fur induktive Abstammungs- und Vererbungslehre, 1928, B. 48, H. 1.
, плодовитость которого обеспечивалась за счет удвоения хромосомных наборов каждого из родительских видов; П. М. Жуковский и А. Р. Жебрак (рис. 207, 208), много сделавшие для изучения полиплоидии у злаков; М. А. Розанова, исследовавшая роль полиплоидии в природе у ягодных форм [386] Розанова M. A. Межвидовая гибридизация в родах Rubus и Fragaria в связи с вопросом о формообразовании // Изв. АН СССР, Сер. биол., 1938, №3, с. 667-679.
; О. С. Стрелкова и А. П. Соколовская, на протяжении десятилетий изучавшие полиплоидию среди цветковых растений в природе и показавшие высокую концентрацию полиплоидов в экстремальных условиях высоких широт и высокогорий [387] Соколовская А. П. Каригеографическое исследование рода Agrostis II Ботан. журн., 1937, т. 22, с. 457-480; Соколовская А. П. Вопросы географического распространения полиплоидных видов растений // Полиплоидия и селекция, М.-Л.: Наука, 1965, с. 1-5-109; Соколовская А. П., Стрелкова О. С. Полиплоидия в высокогорных областях Памира и Алтая // Докл. АН СССР, 1938, т. 21, с. 68-71.
; В. А. Рыбин, осуществивший ресинтез культурной сливы [388] Рыбин В. А. Гибриды терна и алычи и проблема происхождения культурной сливы // Тр. по прикл. ботан., генет. и селекц., 1936, сер. 2, вып. 10; Rybin V. A. Spontane und experimentell erzeugte Bastarde zwischenSchwarzdorn und Kirschtpflaume und das Abstammungsproblem der Kulturpflaume // Planta, 1936, B. 25, S. 22-58/
; Дончо Костов — болгарский генетик, исследовавший полиплоидию у пасленовых, каучуконосов и других растений [389] Костов Дончо. Связь морфологически различных хромосом у межвидовых гибридов Nicotiana // Докл. АН СССР, 1935, №1, с. 555-560; Kostoff D. Polyploids are more variable than their original diploids // Nature, 1939, p. 868-869/
и ресинтезировавший одну из форм табака; В. П. Зосимович, начавший исследование сахарной свеклы с изучения дикорастущих видов рода и полиплоидии у них [390] Зосимович В. П. Межвидовая гибридизация у свекл Beta L. I. Экспериментальный синтез и происхождение Beta tringina Wett. (2n=54) // ДАН СССР, 1938, с. 20-23.
. Не меньший вклад внесли ученые школы Михаила Сергеевича Навашина [391] Навашин М. С. Об изменении числа и морфологических признаков хромосом у межвидовых гибридов // Тр. по прикп. ботанике, генет. и селекции, 1927, т. 17, вып. 3, с. 121-150. Навашин М. С. Проблемы кариологии и цитогенетики в исследованиях на видах рода Crepis. М.:Наука, 1985.
(Е. Н. Герасимова-Навашина, В. А. Поддубная-Арнольди, Н. А. Чуксанова ), а также Лидия Петровна Бреславец [392] Бреславец П. П. Полиплоидия в природе и опыте. М.: Изд-во АН СССР, 1963.
; в Швеции — А. Мюнтцинг [393] Muentzing A. The evolutionary significance of autopolyploidy // Hereditas, 1936, v. 21 d.263-378.
, А. Густафсон [394] Gustafsson A. The plant species in relation to polyploidy and apomixis // Hereditas, 1946, v.32, p.444-448.
, А. Леван [395] Levan A. Zytologische Studien an Allium // Hereditas, 1935, v. 22, p. 1-126.
, А. и Д. Лёвэ [396] Love A., Love D. The signification of differences in the distribution of diploids and polyploids // Hereditas, 1943, v.28, p. 145-163/
, в Японии — X. Кихара и его ученики [397] Kihara H., Nishiyama J. Genomanalyse bei Tnticum und Aegilops. I. // Cytologia (Tolwg), 1930, v.1,p. 270-282.
; в Дании — О. Хагеруп [398] Hagerup O. Dber Polyploidie in Beziehung zu Klima, Okologie und Phylogenie. Chromosomenzahlen aus Timbuktu // Hereditas, 1932, v. 16, p. 19-40; Hagerup О. Studies on the significance of polyploidy // Hereditas, 1938-1940.
, в Германии — Э. Баур [399] Baur E. Artumgrenzung und Artbildung in der Gattung Antirrhinum, Section Antirrinastrum II Zeitsehr fur Vererbungslehre, 1932, B. 58, S. 252-302.
, Г. Тишлер [400] Tischler G. Über die Verwendung der Chromosomenzahl für phylogenetische Probleme bei den Angiospermen // Biol. Zentralblatt, 1928, B.48, S.321-345; Tischler G. Algemeine Pflanzenkaryologie. Ergänzungsband: Angewandte Pflanzenkaryologie. Handbuch der Pflunzenanatomie. Berlin: Nikolassee, 1953.
и Ф. Веттштайн [401] Wettstein von F. Bastardpolyploidie als Artbildungsvorgang bei Pflanzen // Naturwissenschaften, 1932, B.20, S.981-984.
; в Англии — Сирил Дарлингтон (рис. 209) [402] Darlington C. D. Evolution of Genetic Systems. Edinburg: Oliver & Boya, 1939 (переидание—1958).
, создавший учение о хромосомах как генетических системах; большой коллектив американских цитогенетиков и ботаников — Э. Андерсон, Э. Бэбкок, Дж. Болдуин, А. Блексли, А. Эвери, Т. Гудспид [403] Anderson E. The species problem in Iris // Ann. Mont. Botan. Garden, 1936, v.23, p.457-509; Babcock E. B., Cave M. S. A study of intra- and interspecific relations of Crepis foetida L. // Zeitschr. fur induktive Abstammungs- und Vererbungslehre, 1938, B. 75, S. 124-160; Blakeslee A. F., Bergner A. D., Avery A. G. Geographical distribution of chromosomal prime types in Datura stramonium. // Cytologia (Tokyo), 1937, p.1070-1093.
, трио исследователей дикой флоры — Дж. Клаузен, Д. Кек и У. Хизи [404] Clausen J. D., Keck D. D., Hiesey W. M. The concept of species based on experiment // Amer. J. of Botany, 1939, v.26, p.103-106.
, а начиная с 40-х гг. ныне здравствующий выдающийся ботаник-эволюционист Л. Стеббинс (рис. 210) [405] Stebbins L. Variation and Evolution in Plant, N. Y.: Col. Univ. Press, 1950; Stebbins L. Chromosomal Evolution in Higher Plants. London, 1971.
— все они, и десятки других, не упомянутых здесь исследователей на конкретных примерах демонстрировали, в отличие от дрозофилистов, дискретность не только полиплоидии как цитогенетического механизма, но и видообразования в целом. Однако синтетическая теория эволюции (СТЭ) в большей степени создавалась исследователями животных, нежели растений, и этот огромный массив фактов из цитогенетики растений не был в полной мере аккумулирован СТЭ.
Интервал:
Закладка:
Похожие книги на «Развитие эволюционных идей в биологии»
Представляем Вашему вниманию похожие книги на «Развитие эволюционных идей в биологии» списком для выбора. Мы отобрали схожую по названию и смыслу литературу в надежде предоставить читателям больше вариантов отыскать новые, интересные, ещё непрочитанные произведения.












Обсуждение, отзывы о книге «Развитие эволюционных идей в биологии» и просто собственные мнения читателей. Оставьте ваши комментарии, напишите, что Вы думаете о произведении, его смысле или главных героях. Укажите что конкретно понравилось, а что нет, и почему Вы так считаете.