Николай Курчанов - Генетика человека с основами общей генетики. Учебное пособие
Здесь есть возможность читать онлайн «Николай Курчанов - Генетика человека с основами общей генетики. Учебное пособие» — ознакомительный отрывок электронной книги совершенно бесплатно, а после прочтения отрывка купить полную версию. В некоторых случаях можно слушать аудио, скачать через торрент в формате fb2 и присутствует краткое содержание. Город: Санкт-Петербург, Год выпуска: 2009, ISBN: 2009, Издательство: Array Литагент «СпецЛит», Жанр: Биология, Медицина, на русском языке. Описание произведения, (предисловие) а так же отзывы посетителей доступны на портале библиотеки ЛибКат.

- Название:Генетика человека с основами общей генетики. Учебное пособие
- Автор:
- Издательство:Array Литагент «СпецЛит»
- Жанр:
- Год:2009
- Город:Санкт-Петербург
- ISBN:978-5-299-00411-3
- Рейтинг книги:3 / 5. Голосов: 1
-
Избранное:Добавить в избранное
- Отзывы:
-
Ваша оценка:
Генетика человека с основами общей генетики. Учебное пособие: краткое содержание, описание и аннотация
Предлагаем к чтению аннотацию, описание, краткое содержание или предисловие (зависит от того, что написал сам автор книги «Генетика человека с основами общей генетики. Учебное пособие»). Если вы не нашли необходимую информацию о книге — напишите в комментариях, мы постараемся отыскать её.
Второе издание книги значительно переработано автором с учетом новой информации, опубликованной за последние три года.
Пособие предназначено для студентов биологических, педагогических, психологических и социологических факультетов. Представляет интерес для научных работников всех специальностей, занимающихся вопросами, связанными с изучением биологической природы человека.
2-е издание, переработанное и дополненное.
Генетика человека с основами общей генетики. Учебное пособие — читать онлайн ознакомительный отрывок
Ниже представлен текст книги, разбитый по страницам. Система сохранения места последней прочитанной страницы, позволяет с удобством читать онлайн бесплатно книгу «Генетика человека с основами общей генетики. Учебное пособие», без необходимости каждый раз заново искать на чём Вы остановились. Поставьте закладку, и сможете в любой момент перейти на страницу, на которой закончили чтение.
Интервал:
Закладка:
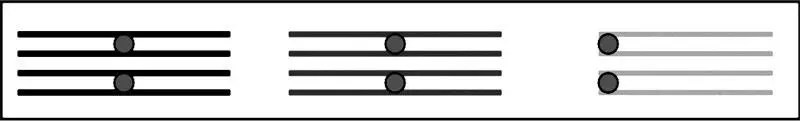
Рис. 3.4. Синапсис гомологичных хромосом с образованием бивалентов в профазе мейоза
Профаза-1. Наиболее сложная, длительная и важная стадия мейоза. Помимо процессов, аналогичных процессам профазы митоза (спирализация хромосом, разрушение ядерной мембраны, исчезновение ядрышка, образование веретена деления), определяющее значение для всего последующего процесса имеет конъюгация гомологичных хромосом – синапсис.Соединенные пары гомологов называются бивалентами(рис. 3.4).
Гомологичные хромосомы связывает особая структура, образованная из белков кариоплазмы – синаптонемный комплекс (СК).В бивалентах гомологичные хромосомы могут обмениваться гомологичными участками. Такой процесс называется кроссинговером.Механизм кроссинговера довольно сложен. Кроссинговер вносит большой вклад в повышение генетического разнообразия, играет важную эволюционную роль и активно изучается на протяжении всей истории генетики. Однако до сих пор он сохраняет свои загадки.
В связи с длительностью и многообразием процессов профазы-1 ее обычно подразделяют на 5 подстадий.
Лептотена – начало спирализации и уплотнения хромосом.
Зиготена – начало (с отдельных участков) и завершение синапсиса гомологичных хромосом. Происходит формирование СК.
Пахитена – укорочение и утолщение бивалентов (стадия толстых нитей).
Диплотена – гомологичные хромосомы бивалентов начинают расходиться (разрушается СК), но они связаны в нескольких зонах контакта – хиазмах.Число хиазм в биваленте может быть различным (обычно 2–3), в длинных хромосомах больше, чем в коротких. Хиазмычасто показывают, что между хроматидами происходит кроссинговер.
Диакинез – хромосомы достигают максимальной спирализации. Исчезают хиазмы, и к концу диакинеза хромосомы остаются связанными только в теломерных участках.
В конце профазы-1 центриоли расходятся к полюсам клетки.
Метафаза-1. Завершается формирование веретена деления. Биваленты концентрируются в экваториальной плоскости клетки.
Анафаза-1. Гомологичные хромосомы расходятся к полюсам клетки. Каждая хромосома состоит из двух хроматид, соединенных общей центромерой.
Телофаза-1. Обычно очень короткая. У полюсов клетки группируются гаплоидные наборы хромосом, в которых представлен только один из парыгомологов. Восстанавливаются структура ядра и ядерная мембрана. Происходит частичная деспирализация хромосом. В конце телофазы-1 наступает цитокинез и образуются две клетки с гаплоидным набором хромосом.
После телофазы-1 вновь образованные клетки сразу вступают в мейоз-2, который проходит по типу обычного митоза.
Профаза-2. Частично деспирализованные хромосомы хорошо различимы. Начинается процесс обратной спирализации хромосом. Разрушается ядерная мембрана, формируется веретено деления, центриоли начинают расходиться к полюсам клетки.
Метафаза-2. Хромосомы выстраиваются в экваториальной плоскости. Центромеры прикрепляются к микротрубочкам образованного веретена деления.
Анафаза-2. Происходит разделение центромер, и каждая хроматида становится самостоятельной хромосомой. Дочерние хромосомы направляются к полюсам клетки.
Телофаза-2. Формируются новые ядра с гаплоидным набором хромосом. Хромосомы деконденсируются. Наступает цитокинез.
Основное биологическое значение мейоза заключается в обеспечении постоянства числа хромосом на протяжении поколений при половом размножении. Важным следствием мейоза является обеспечение генетического разнообразия гамет в результате рекомбинации хромосом и кроссинговера.
Механизм распределения неядерных генетических структур (митохондрий, хлоропластов) при митозе и мейозе пока неизвестен.
Глава 4. Закономерности наследственности
Ключевой проблемой биологии, по-видимому, можно считать вопрос о том, как увековечивает свой опыт живая материя.
М. Дельбрюк (1906–1981), американский генетик, лауреат Нобелевской премии 1969 г.Общебиологическое значение генетики обусловлено тем, что законы наследственности справедливы для всех организмов. Понятия, сформировавшиеся при изучении закономерностей наследования, являются базовыми для всех разделов генетики.
Читать дальшеИнтервал:
Закладка:
Похожие книги на «Генетика человека с основами общей генетики. Учебное пособие»
Представляем Вашему вниманию похожие книги на «Генетика человека с основами общей генетики. Учебное пособие» списком для выбора. Мы отобрали схожую по названию и смыслу литературу в надежде предоставить читателям больше вариантов отыскать новые, интересные, ещё непрочитанные произведения.












Обсуждение, отзывы о книге «Генетика человека с основами общей генетики. Учебное пособие» и просто собственные мнения читателей. Оставьте ваши комментарии, напишите, что Вы думаете о произведении, его смысле или главных героях. Укажите что конкретно понравилось, а что нет, и почему Вы так считаете.