Максим Франк-Каменецкий - Самая главная молекула. От структуры ДНК к биомедицине XXI века
Здесь есть возможность читать онлайн «Максим Франк-Каменецкий - Самая главная молекула. От структуры ДНК к биомедицине XXI века» — ознакомительный отрывок электронной книги совершенно бесплатно, а после прочтения отрывка купить полную версию. В некоторых случаях можно слушать аудио, скачать через торрент в формате fb2 и присутствует краткое содержание. Город: Москва, Год выпуска: 2017, ISBN: 2017, Издательство: Литагент Альпина, Жанр: Биология, Биология, Прочая научная литература, на русском языке. Описание произведения, (предисловие) а так же отзывы посетителей доступны на портале библиотеки ЛибКат.

- Название:Самая главная молекула. От структуры ДНК к биомедицине XXI века
- Автор:
- Издательство:Литагент Альпина
- Жанр:
- Год:2017
- Город:Москва
- ISBN:978-5-9614-4522-0
- Рейтинг книги:3 / 5. Голосов: 1
-
Избранное:Добавить в избранное
- Отзывы:
-
Ваша оценка:
Самая главная молекула. От структуры ДНК к биомедицине XXI века: краткое содержание, описание и аннотация
Предлагаем к чтению аннотацию, описание, краткое содержание или предисловие (зависит от того, что написал сам автор книги «Самая главная молекула. От структуры ДНК к биомедицине XXI века»). Если вы не нашли необходимую информацию о книге — напишите в комментариях, мы постараемся отыскать её.
Самая главная молекула. От структуры ДНК к биомедицине XXI века — читать онлайн ознакомительный отрывок
Ниже представлен текст книги, разбитый по страницам. Система сохранения места последней прочитанной страницы, позволяет с удобством читать онлайн бесплатно книгу «Самая главная молекула. От структуры ДНК к биомедицине XXI века», без необходимости каждый раз заново искать на чём Вы остановились. Поставьте закладку, и сможете в любой момент перейти на страницу, на которой закончили чтение.
Интервал:
Закладка:
Давайте оценим, последовательность какой длины такой инструмент должен узнавать. Будем считать для простоты, что геномная ДНК представляет собой чисто случайную последовательность четырех нуклеотидов А, Т, Г, Ц, причем все четыре нуклеотида встречаются с одинаковой вероятностью (это, конечно, грубое предположение, но для приблизительной оценки вполне годится). Тогда вероятность встретить конкретную последовательность из n нуклеотидов будет, очевидно, 4 — n . Такая последовательность встретится в геноме, состоящем из N нуклеотидов, N 4 — n раз. Следовательно, чтобы последовательность встретилась всего один раз, нужно, чтобы выполнялось условие: N 4 — n = 1. Из этого уравнения легко находим: n = log N / log4 – и, вспомнив, что человеческий геном состоит из 310 9нуклеотидов, получаем для n значение 16. Таким образом, для того, чтобы последовательность не повторялась в геноме, т. е. была уникальной, она должна состоять не менее чем из 16 нуклеотидов. Теперь мы видим, насколько рестриктазы – негодный инструмент для редактирования генома.
Химики, биохимики и биофизики взялись за поиск адекватного инструмента, в частности, автор этих строк приложил большие усилия в этом направлении. Первой идеей было использовать способность ДНК образовывать тройную спираль, о чем мы уже говорили в главе 9. Идея казалась очень привлекательной. Достаточно выбрать целевую последовательность в геноме из, скажем, 16 нуклеотидов и синтезировать соответствующую цепь однонитевой ДНК из 16 нуклеотидов, которая образовывала бы с выбранным участком геномной ДНК тройную спираль. На один из концов синтетической ДНК можно приладить активную химическую группу или даже целый белок-эндонуклеазу, способный разрезать ДНК там, куда он доставлен. Но проблема с триплексами в том, что они образуются только в тех местах генома, в которых в одной цепи стоят одни пурины (А и Г), а, соответственно, в комплементарной цепи одни пиримидины (Т и Ц). Такие участки длиной в 16 или более нуклеотидов редко встречаются в геноме, что резко сужает выбор целевых участков для разрезания. Так что от ДНКовых триплексов пришлось отказаться.
Следующей идеей было использовать синтетический аналог ДНК, ПНК, или пептидную нуклеиновую кислоту. Этот очень интересный синтетический аналог ДНК был изобретен в 1991 году в группе Питера Нильсена в университете Копенгагена. ПНК имеет те же основания, что и ДНК, но вместо сахарофосфатного остова основания присоединены к пептидному остову, похожему на остов белковой цепи (рис. 45). Поскольку, в отличие от ДНК, ПНК не несет отрицательного заряда, две молекулы ПНК образуют с одиночной цепью ДНК очень прочные триплексы. Эти триплексы настолько прочные, что две молекулы ПНК в определенных условиях способны раскрыть двойную спираль, образовав триплекс с одной из цепей, оставив комплементарную цепь без партнера (рис. 45). У ПНК множество применений, но нас сейчас интересует одно из них, состоящее в превращении обычных рестриктаз, узнающих последовательности из шести нуклеотидов, в инструмент с гораздо большей избирательностью. Как это делается, схематически показано на рис. 45.
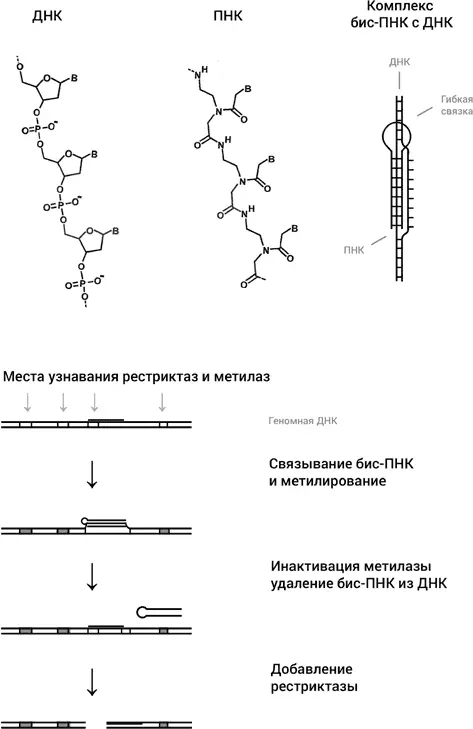
Рис. 45.ПНК имеет те же основания, что и ДНК (обозначены как В), но они прикреплены к совсем другому остову, чем в ДНК, напоминающему остов молекулы белка. Так называемая бис-ПНК, состоящая из двух коротких молекул ПНК, связанных гибкой молекулярной связкой, образует триплекс с одной из цепей ДНК (посредством триад, изображенных на рис. 42), оставляя комплементарную цепь в виде однонитевой петли. В нижней части рисунка дана схема того, как бис-ПНК используется для того, чтобы превратить обычную рестриктазу в очень редко расщепляющий ДНК инструмент
Целевой участок в геноме для связывания двух молекул ПНК с образованием триплекса выбирается таким образом, чтобы он чуть-чуть перекрывался с участком связывания какой-нибудь рестриктазы. После связывания ПНК с ДНК препарат обрабатывается соответствующей рестриктазе метилазой. Метилаза метилирует все участки узнавания в геноме, кроме одного, который стал недоступен для связывания метилазы, так как его дуплексная структура была нарушена связыванием ПНК. Затем делается так, чтобы связывание ПНК с ДНК было разрушено. В результате единственное место связывания рестриктазы с ДНК восстанавливается, тогда как все остальные места связывания не работают, поскольку они прометилированы. Теперь рестриктаза разрежет ДНК только в одном месте, в том, которое было изначально выбрано для этой цели. Конечно, мы несколько утрировали ситуацию, неметилированных участков в геноме может оказаться несколько, но ясно, что такой подход должен резко увеличивать избирательность нуклеаз, что и было экспериментально продемонстрировано в моей лаборатории в Бостонском университете в работе, выполненной совместно с Нильсеном.
Читать дальшеИнтервал:
Закладка:
Похожие книги на «Самая главная молекула. От структуры ДНК к биомедицине XXI века»
Представляем Вашему вниманию похожие книги на «Самая главная молекула. От структуры ДНК к биомедицине XXI века» списком для выбора. Мы отобрали схожую по названию и смыслу литературу в надежде предоставить читателям больше вариантов отыскать новые, интересные, ещё непрочитанные произведения.












Обсуждение, отзывы о книге «Самая главная молекула. От структуры ДНК к биомедицине XXI века» и просто собственные мнения читателей. Оставьте ваши комментарии, напишите, что Вы думаете о произведении, его смысле или главных героях. Укажите что конкретно понравилось, а что нет, и почему Вы так считаете.