Максим Франк-Каменецкий - Самая главная молекула. От структуры ДНК к биомедицине XXI века
Здесь есть возможность читать онлайн «Максим Франк-Каменецкий - Самая главная молекула. От структуры ДНК к биомедицине XXI века» — ознакомительный отрывок электронной книги совершенно бесплатно, а после прочтения отрывка купить полную версию. В некоторых случаях можно слушать аудио, скачать через торрент в формате fb2 и присутствует краткое содержание. Город: Москва, Год выпуска: 2017, ISBN: 2017, Издательство: Литагент Альпина, Жанр: Биология, Биология, Прочая научная литература, на русском языке. Описание произведения, (предисловие) а так же отзывы посетителей доступны на портале библиотеки ЛибКат.

- Название:Самая главная молекула. От структуры ДНК к биомедицине XXI века
- Автор:
- Издательство:Литагент Альпина
- Жанр:
- Год:2017
- Город:Москва
- ISBN:978-5-9614-4522-0
- Рейтинг книги:3 / 5. Голосов: 1
-
Избранное:Добавить в избранное
- Отзывы:
-
Ваша оценка:
Самая главная молекула. От структуры ДНК к биомедицине XXI века: краткое содержание, описание и аннотация
Предлагаем к чтению аннотацию, описание, краткое содержание или предисловие (зависит от того, что написал сам автор книги «Самая главная молекула. От структуры ДНК к биомедицине XXI века»). Если вы не нашли необходимую информацию о книге — напишите в комментариях, мы постараемся отыскать её.
Самая главная молекула. От структуры ДНК к биомедицине XXI века — читать онлайн ознакомительный отрывок
Ниже представлен текст книги, разбитый по страницам. Система сохранения места последней прочитанной страницы, позволяет с удобством читать онлайн бесплатно книгу «Самая главная молекула. От структуры ДНК к биомедицине XXI века», без необходимости каждый раз заново искать на чём Вы остановились. Поставьте закладку, и сможете в любой момент перейти на страницу, на которой закончили чтение.
Интервал:
Закладка:
Все же подход, основанный на ПНК, оказался слишком сложным, и, что самое главное, его не удавалось использовать в живой клетке. А именно редактирование ДНК непосредственно в живой клетке представляет наибольший интерес. К началу 2010 года два очень изощренных биохимические метода специфического разрезания ДНК были разработаны в биотехнологических компаниях. И их уже стали применять для редактирования геномов. Но в начале 2013 года произошел подлинный прорыв, который самым радикальным образом изменил ситуацию. Появился метод, носящий неуклюжее название КРИСПР-кас.
Метод целиком базируется на системе приобретенного иммунитета у бактерий, о котором рассказано в главе 6.
Для редактирования генома действующие лица иммунной защиты бактерий, крРНК и кас-белок, переносят в эукариотическую клетку, при этом дизайн молекулы крРНК делается с таким расчетом, чтобы промежуточная последовательность в ней была идентична выбранному участку в геномной ДНК эукариотической клетки. «Как это можно?! – слышу изумленный возглас читателя. – Система произведет двунитевой разрыв в ДНК, на этом все и закончится: клетка погибнет». Нет, не закончится. Ведь недаром мы диплоидные существа: у каждого участка нашей аутосомной ДНК есть двойник, гомологичный участок, расположенный на сестринской хромосоме. Наличие гомологичного участка позволяет нашим соматическим клеткам залечивать или, как говорят, репарировать ДНК, которая подверглась самому опасному повреждению: двунитевому разрыву. Этот механизм репарации двунитевых разрывов называется гомологичной рекомбинацией. В деталях гомологичная рекомбинация – это сложный процесс, и он происходит не только при репарации, но и в других случаях.
Для нас важно следующее: для того чтобы гомологичная рекомбинация репарировала двунитевой разрыв, необходимо, чтобы в клетки находилась ДНК с двумя последовательностями, идентичными двум последовательностям, расположенным справа и слава от места разрыва. Бактериальная ДНК гораздо более уязвима в отношении двунитевых разрывов именно потому, что бактерии – это гаплоидные существа, у них имеется только одна-единственная копия генома, и поэтому никакая гомологичная рекомбинация невозможна. Именно гомологическая рекомбинация не только спасает эукариотическую клетку от гибели, когда в клетку вводят крРНК и кас-белок, но и позволяет осуществить редактирование генома. Как это делается?
Давайте разберем ситуацию на конкретном примере. Представим себе, что мы решили осуществить генную терапию больного серповидно-клеточной анемией (СКА), о которой шла речь в главе 2 (и еще пойдет в главе 12). Иными словами, мы хотим заменить мутантный ген у больного на здоровый ген β-цепи гемоглобина, не несущий мутацию СКА (рис. 46). Используя генную инженерию, мы готовим вектор, которым может быть плазмида или ДНК обезвреженного аденовируса. В вектор мы встраиваем следующие добавочные участки: два участка, кодирующие две молекулы крРНК, причем дизайн одной сделан так, чтобы она вызывала разрыв около левого края гена СКА, а дизайн другой сделан так, чтобы разрыв был около правого края; ген, кодирующий кас-белок. Мы размножаем взятые из пациента клетки – предшественницы красных кровяных клеток и вводим в них наш вектор, а также отрезок двунитевой ДНК, состоящий из здорового гена β-цепи гемоглобина, не несущего мутации СКА, слева и справа от которого имеются точно такие же последовательности (они обозначены как ГЛ и ГП на рис. 46), как последовательности слева и справа от двунитевого разрыва в геномной ДНК, который получится после удаления из генома гена СКА с помощью двух молекул крРНК, которые закодированы в векторе.
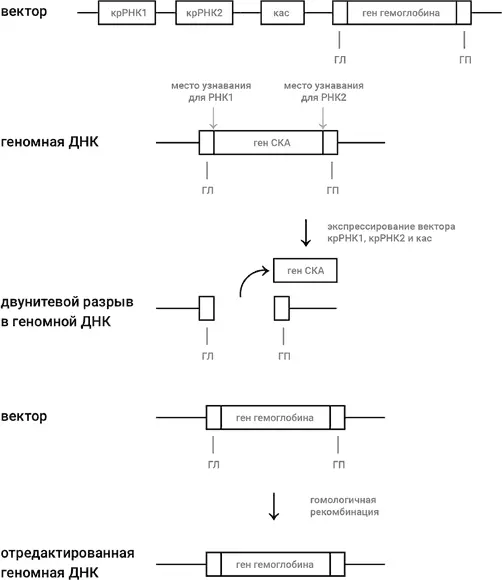
Рис. 46.Редактирование генома при помощи технологии КРИСПР-кас. Стандартными методами генной инженерии создается вектор (плазмида или обезвреженный аденовирус), в который включены гены, кодирующие две крРНК, узнающие левый и правый концы гена, который будет заменен (в нашем случае это ген СКА), ген кас и ген, на который мы хотим заменить ген СКА, т. е. в нашем случае – нормальный ген β-цепи гемоглобина. Слева и справа от гена β-цепи гемоглобина должны быть участки, гомологичные (т. е. идентичные) тем участкам, которые соседствуют с ним (а также с геном СКА) в геноме (они обозначены как ГЛ и ГП). Введение – вектора в клетку – предшественницу красных кровяных шариков приводит к экспрессии крРНК1, крРНК2 и кас-белка, что в свою очередь приводит к выбрасыванию гена СКА из генома и к двунитевому разрыву в ДНК, как было объяснено в главе 6 (см. рис. 25). В клетке – предшественнице красных кровяных шариков включается система репарации двунитевого разрыва, называемая «гомологичной рекомбинацией». Наличие участков ГЛ и ГП в векторе (или в отдельно введенном отрезке ДНК, несущем здоровый ген) приводит к тому, что ген β-цепи гемоглобина переносится из вектора в образовавшуюся брешь – замена мутантного гена СКА на здоровый ген β-цепи гемоглобина совершена
Читать дальшеИнтервал:
Закладка:
Похожие книги на «Самая главная молекула. От структуры ДНК к биомедицине XXI века»
Представляем Вашему вниманию похожие книги на «Самая главная молекула. От структуры ДНК к биомедицине XXI века» списком для выбора. Мы отобрали схожую по названию и смыслу литературу в надежде предоставить читателям больше вариантов отыскать новые, интересные, ещё непрочитанные произведения.












Обсуждение, отзывы о книге «Самая главная молекула. От структуры ДНК к биомедицине XXI века» и просто собственные мнения читателей. Оставьте ваши комментарии, напишите, что Вы думаете о произведении, его смысле или главных героях. Укажите что конкретно понравилось, а что нет, и почему Вы так считаете.