Александр Марков - Перспективы отбора. От зеленых пеночек и бессмысленного усложнения до голых землекопов и мутирующего человечества
Здесь есть возможность читать онлайн «Александр Марков - Перспективы отбора. От зеленых пеночек и бессмысленного усложнения до голых землекопов и мутирующего человечества» — ознакомительный отрывок электронной книги совершенно бесплатно, а после прочтения отрывка купить полную версию. В некоторых случаях можно слушать аудио, скачать через торрент в формате fb2 и присутствует краткое содержание. Город: Москва, Год выпуска: 2019, ISBN: 2019, Издательство: АСТ, Corpus, Жанр: Биология, sci_popular, на русском языке. Описание произведения, (предисловие) а так же отзывы посетителей доступны на портале библиотеки ЛибКат.
- Название:Перспективы отбора. От зеленых пеночек и бессмысленного усложнения до голых землекопов и мутирующего человечества
- Автор:
- Издательство:АСТ, Corpus
- Жанр:
- Год:2019
- Город:Москва
- ISBN:978-5-17-114115-8
- Рейтинг книги:4 / 5. Голосов: 1
-
Избранное:Добавить в избранное
- Отзывы:
-
Ваша оценка:
Перспективы отбора. От зеленых пеночек и бессмысленного усложнения до голых землекопов и мутирующего человечества: краткое содержание, описание и аннотация
Предлагаем к чтению аннотацию, описание, краткое содержание или предисловие (зависит от того, что написал сам автор книги «Перспективы отбора. От зеленых пеночек и бессмысленного усложнения до голых землекопов и мутирующего человечества»). Если вы не нашли необходимую информацию о книге — напишите в комментариях, мы постараемся отыскать её.
Перспективы отбора. От зеленых пеночек и бессмысленного усложнения до голых землекопов и мутирующего человечества — читать онлайн ознакомительный отрывок
Ниже представлен текст книги, разбитый по страницам. Система сохранения места последней прочитанной страницы, позволяет с удобством читать онлайн бесплатно книгу «Перспективы отбора. От зеленых пеночек и бессмысленного усложнения до голых землекопов и мутирующего человечества», без необходимости каждый раз заново искать на чём Вы остановились. Поставьте закладку, и сможете в любой момент перейти на страницу, на которой закончили чтение.
Интервал:
Закладка:
Проблема, однако, в том, что найти подходящих для такого анализа ископаемых животных очень непросто. Во-первых, их «захоронения» должны быть массовыми, а ископаемая летопись — подробной, чтобы моменты появления и вымирания видов поддавались точному датированию. Во-вторых, у этих животных должен быть четкий половой диморфизм, по выраженности которого можно судить о силе полового отбора. Причем диморфизм должен затрагивать не только мягкие ткани, крайне редко сохраняющиеся в ископаемом состоянии, но и скелет. Желательно также, чтобы группа не была полностью вымершей (как, например, аммониты) и имела живых представителей, которые позволили бы убедиться, что различия, принимаемые нами за межполовые, действительно таковы (а не межвидовые или, скажем, возрастные).
Чуть ли не единственной группой, удовлетворяющей этим критериям, являются остракоды, или ракушковые, — вездесущие мелкие ракообразные, населяющие моря и пресные воды с кембрийского периода по настоящее время. Тело остракод заключено в прочную двустворчатую раковину, отлично сохраняющуюся в ископаемом состоянии. При этом у многих остракод хорошо выражен половой диморфизм (рис. 13.1).
У современных представителей цитероидных остракод (надсемейство Cytheroidea) раковинки самцов сильнее вытянуты в передне-заднем направлении. Это связано с тем, что в задней части раковины у самца помещается громоздкий копулятивный аппарат. Хотя сам аппарат не сохраняется в ископаемом состоянии, форма створок недвусмысленно свидетельствует о его наличии или отсутствии, что позволяет уверенно отличать самцов от самок на ископаемом материале (рис. 13.2).
У современных цитероидных остракод размер мужских половых органов не связан с какими-либо особенностями брачного поведения (ухаживанием или сигналами, подаваемыми самке), а связан лишь с количеством производимой спермы и силой мышц, которые эту сперму выбрасывают во время спаривания. По-видимому, гипертрофия половых органов у самцов является результатом спермовых войн.
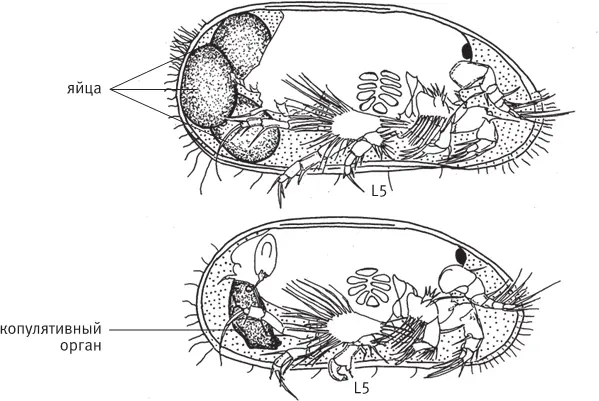
рис. 13.1.Половой диморфизм у современной остракоды Vestalenula cornelia . Вид справа, правая створка удалена. Вверху — самка, внизу — самец с крупным копулятивным органом. Половой диморфизм затрагивает также строение пятой пары конечностей ( L5 ), которую самцы используют для удержания самки во время копуляции. По рисунку из Ozawa, 2013 .
Если самцу приходится тратить ресурсы на выращивание огромных половых органов, то у него, при прочих равных условиях, должно оставаться меньше сил на все остальное. Спермовые войны предполагают острую конкуренцию между самцами, что может вредить здоровью самок. В таком случае самки в ответ могут выработать защитные адаптации, тоже затратные. По остракодам таких данных нет, но это следует из общих соображений и показано на других членистоногих (см. Исследование № 15). Меняя морфологию и физиологию самцов и самок, сильный половой отбор, очевидно, уводит их от того оптимального строения, к которому привел бы их обычный естественный отбор в отсутствие полового. Иными словами, организмы приобретают строение, неоптимальное с точки зрения выживаемости и способности переносить невзгоды, но связанное с взаимоотношениями между полами. Поэтому логично предположить, что виды остракод с сильным половым диморфизмом будут вымирать в среднем чаще, чем виды со слабым диморфизмом.
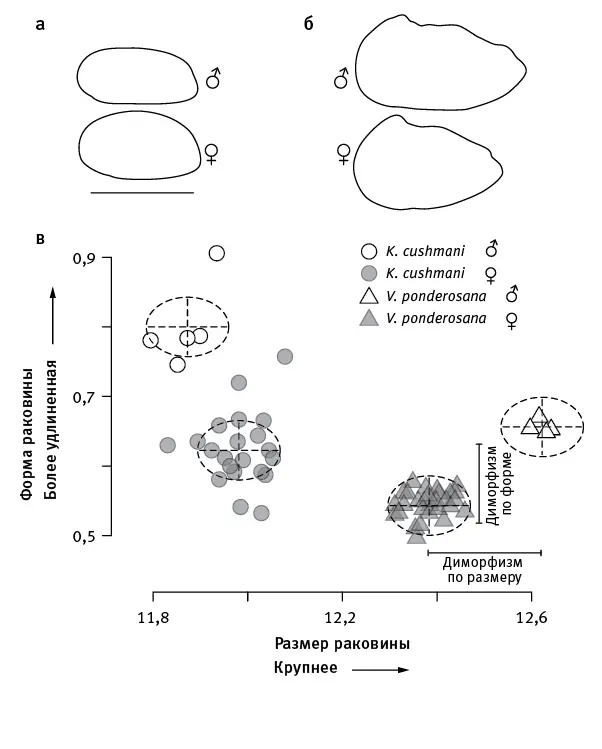
рис. 13.2.Половой диморфизм у двух видов позднемеловых цитероидных остракод. а — очертания раковин самцов и самок Krithe cushmani, б — Veenia ponderosana . На графике ( в ) по вертикальной оси отложена степень удлиненности раковины (логарифм отношения длины к высоте), по горизонтальной — размер раковины (логарифм площади створки). Видно, что у обоих видов самцы имеют более удлиненную раковину, чем самки (это справедливо и для всех остальных цитероидных остракод). Что касается диморфизма по размеру, то он может иметь разную направленность: у K. cushmani самцы мельче самок, у V. ponderosana — крупнее. По рисунку из Martins et al., 2018 .
Для проверки этого предположения американские палеонтологи проанализировали подробнейшие данные по распространению 93 видов цитероидных остракод в верхнемеловых отложениях юго-востока США, штаты Миссисипи и Алабама ( Martins et al., 2018). Возраст изученных слоев — от 66 до 84 млн лет. Исследованные виды сильно различаются по выраженности полового диморфизма.
Читать дальшеИнтервал:
Закладка:
Похожие книги на «Перспективы отбора. От зеленых пеночек и бессмысленного усложнения до голых землекопов и мутирующего человечества»
Представляем Вашему вниманию похожие книги на «Перспективы отбора. От зеленых пеночек и бессмысленного усложнения до голых землекопов и мутирующего человечества» списком для выбора. Мы отобрали схожую по названию и смыслу литературу в надежде предоставить читателям больше вариантов отыскать новые, интересные, ещё непрочитанные произведения.










Обсуждение, отзывы о книге «Перспективы отбора. От зеленых пеночек и бессмысленного усложнения до голых землекопов и мутирующего человечества» и просто собственные мнения читателей. Оставьте ваши комментарии, напишите, что Вы думаете о произведении, его смысле или главных героях. Укажите что конкретно понравилось, а что нет, и почему Вы так считаете.