Александр Марков - Перспективы отбора. От зеленых пеночек и бессмысленного усложнения до голых землекопов и мутирующего человечества
Здесь есть возможность читать онлайн «Александр Марков - Перспективы отбора. От зеленых пеночек и бессмысленного усложнения до голых землекопов и мутирующего человечества» — ознакомительный отрывок электронной книги совершенно бесплатно, а после прочтения отрывка купить полную версию. В некоторых случаях можно слушать аудио, скачать через торрент в формате fb2 и присутствует краткое содержание. Город: Москва, Год выпуска: 2019, ISBN: 2019, Издательство: АСТ, Corpus, Жанр: Биология, sci_popular, на русском языке. Описание произведения, (предисловие) а так же отзывы посетителей доступны на портале библиотеки ЛибКат.
- Название:Перспективы отбора. От зеленых пеночек и бессмысленного усложнения до голых землекопов и мутирующего человечества
- Автор:
- Издательство:АСТ, Corpus
- Жанр:
- Год:2019
- Город:Москва
- ISBN:978-5-17-114115-8
- Рейтинг книги:4 / 5. Голосов: 1
-
Избранное:Добавить в избранное
- Отзывы:
-
Ваша оценка:
Перспективы отбора. От зеленых пеночек и бессмысленного усложнения до голых землекопов и мутирующего человечества: краткое содержание, описание и аннотация
Предлагаем к чтению аннотацию, описание, краткое содержание или предисловие (зависит от того, что написал сам автор книги «Перспективы отбора. От зеленых пеночек и бессмысленного усложнения до голых землекопов и мутирующего человечества»). Если вы не нашли необходимую информацию о книге — напишите в комментариях, мы постараемся отыскать её.
Перспективы отбора. От зеленых пеночек и бессмысленного усложнения до голых землекопов и мутирующего человечества — читать онлайн ознакомительный отрывок
Ниже представлен текст книги, разбитый по страницам. Система сохранения места последней прочитанной страницы, позволяет с удобством читать онлайн бесплатно книгу «Перспективы отбора. От зеленых пеночек и бессмысленного усложнения до голых землекопов и мутирующего человечества», без необходимости каждый раз заново искать на чём Вы остановились. Поставьте закладку, и сможете в любой момент перейти на страницу, на которой закончили чтение.
Интервал:
Закладка:
Кроме того, в бесполой популяции полезные мутации, возникшие в разных генах у разных особей, не могут объединиться. Вместо этого их носители начинают конкурировать и вытеснять друг друга, что ведет к безвозвратной потере большинства полезных мутаций, возникающих в популяции. Это называют конкуренцией клонов, или клональной интерференцией (см. Исследование № 4).
Все это очень логично, но для полной уверенности не хватает прямых молекулярно-генетических данных об эволюционных изменениях в половых и бесполых популяциях. Чтобы их получить, нужно, во-первых, создать «правильные» подопытные популяции, не отличающиеся друг от друга ничем, кроме наличия или отсутствия полового размножения, а во-вторых — проследить за изменениями их геномов в ходе эволюции.
Именно это и сделали недавно биологи из Гарвардского университета (США). Они провели эволюционный эксперимент на специально созданных для этой цели штаммах пекарских дрожжей, Saccharomyces cerevisiae ( McDonald et al., 2016). В эксперименте участвовало 18 популяций: 12 бесполых, размножавшихся только почкованием, и 6 половых, которые тоже бóльшую часть времени размножались почкованием, но после каждых 90 бесполых поколений их заставляли спариваться и производить потомство половым путем. Все популяции были помещены в одинаковые условия, к которым они приспосабливались в ходе эксперимента, продолжавшегося 100 дней (примерно 1000 поколений).
Жизненный цикл дрожжей изображен на рис. 7.1. Гаплоидные клетки делятся на два пола: a и α. Они могут размножаться почкованием, а могут слиться и превратиться в диплоидную клетку. Диплоидные дрожжи тоже могут размножаться почкованием, а могут в результате мейоза превратиться в четыре гаплоидные споры, две из которых будут принадлежать к полу a , а две — к α.
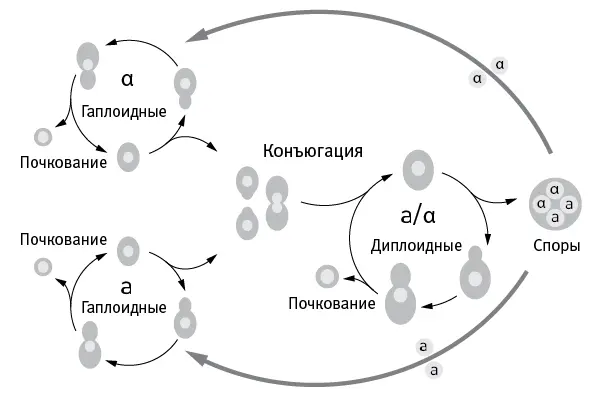
рис. 7.1.Жизненный цикл пекарских дрожжей ( Saccharomyces cerevisiae ).
В ходе эксперимента клетки бесполых популяций всегда находились в гаплоидной фазе и размножались исключительно почкованием. Чтобы они не спаривались, каждая популяция состояла из клеток только одного пола (шесть популяций a и шесть α).
Каждая половая популяция состояла из двух разнополых половинок, которые содержались раздельно в течение 90 поколений, а затем смешивались и помещались в условия, стимулирующие половой процесс. После этого отбирались диплоидные клетки, возникшие в результате спаривания, и помещались в условия, способствующие образованию спор. Затем отбраковывались диплоидные клетки, не захотевшие превращаться в споры, а гаплоидные клетки, полученные из спор, разделялись по половому признаку и снова содержались раздельно в течение следующих 90 поколений.
Чтобы провести все эти манипуляции, нужно иметь возможность быстро и эффективно отделять диплоидные клетки от гаплоидных, а также пол a от α. Для этого в геномы дрожжей были внесены изменения. В частности, были добавлены гены устойчивости к двум антибиотикам, причем так, чтобы клетки a оказались устойчивы только к первому антибиотику, клетки α — только ко второму, а диплоидные клетки — к обоим. Благодаря другим модификациям только гаплоидные клетки пола a умели синтезировать гистидин, лишь гаплоидные клетки пола α могли производить лейцин, а синтез урацила могли осуществлять гаплоидные клетки обоих полов, но не диплоидные. В результате исследователи получили возможность легко (в работе биологов-экспериментаторов все относительно, в том числе и что значит «легко») отбирать нужные клетки и контролировать чистоту опыта, попеременно используя питательные среды, содержащие (или не содержащие) урацил, гистидин, лейцин и два антибиотика в разных комбинациях.
Условия содержания бесполых и половых популяций сделали настолько одинаковыми, насколько это было возможно. Численность всех популяций поддерживалась примерно на одном уровне (порядка 100 000 особей), они содержались в одинаковых емкостях, при одинаковой температуре и на одинаковой питательной среде.
По прошествии 1000 поколений была измерена приспособленность подопытных популяций. Для этого предковый штамм, помеченный флуоресцентной меткой, смешивали в равной пропорции с тестируемым штаммом и выращивали в течение 30 поколений в тех же условиях, в которых проходил основной эксперимент, а потом подсчитывали долю меченых клеток. Чем она меньше, тем выше приспособленность тестируемого штамма. Оказалось, что половые популяции адаптировались лучше бесполых: у первых приспособленность за 1000 поколений выросла на 10–15 %, у вторых — лишь на 5–10 %. Но это результат, можно сказать, тривиальный: способность полового размножения ускорять адаптацию уже не раз подтверждалась в экспериментах (о некоторых из них рассказано в нашей книге «Эволюция. Классические идеи в свете новых открытий»). Хотелось бы знать, что конкретно происходит с геномами половых и бесполых особей и каков генетический базис повышенного темпа адаптации половых популяций.
Читать дальшеИнтервал:
Закладка:
Похожие книги на «Перспективы отбора. От зеленых пеночек и бессмысленного усложнения до голых землекопов и мутирующего человечества»
Представляем Вашему вниманию похожие книги на «Перспективы отбора. От зеленых пеночек и бессмысленного усложнения до голых землекопов и мутирующего человечества» списком для выбора. Мы отобрали схожую по названию и смыслу литературу в надежде предоставить читателям больше вариантов отыскать новые, интересные, ещё непрочитанные произведения.










Обсуждение, отзывы о книге «Перспективы отбора. От зеленых пеночек и бессмысленного усложнения до голых землекопов и мутирующего человечества» и просто собственные мнения читателей. Оставьте ваши комментарии, напишите, что Вы думаете о произведении, его смысле или главных героях. Укажите что конкретно понравилось, а что нет, и почему Вы так считаете.