Александр Марков - Перспективы отбора. От зеленых пеночек и бессмысленного усложнения до голых землекопов и мутирующего человечества
Здесь есть возможность читать онлайн «Александр Марков - Перспективы отбора. От зеленых пеночек и бессмысленного усложнения до голых землекопов и мутирующего человечества» — ознакомительный отрывок электронной книги совершенно бесплатно, а после прочтения отрывка купить полную версию. В некоторых случаях можно слушать аудио, скачать через торрент в формате fb2 и присутствует краткое содержание. Город: Москва, Год выпуска: 2019, ISBN: 2019, Издательство: АСТ, Corpus, Жанр: Биология, sci_popular, на русском языке. Описание произведения, (предисловие) а так же отзывы посетителей доступны на портале библиотеки ЛибКат.
- Название:Перспективы отбора. От зеленых пеночек и бессмысленного усложнения до голых землекопов и мутирующего человечества
- Автор:
- Издательство:АСТ, Corpus
- Жанр:
- Год:2019
- Город:Москва
- ISBN:978-5-17-114115-8
- Рейтинг книги:4 / 5. Голосов: 1
-
Избранное:Добавить в избранное
- Отзывы:
-
Ваша оценка:
Перспективы отбора. От зеленых пеночек и бессмысленного усложнения до голых землекопов и мутирующего человечества: краткое содержание, описание и аннотация
Предлагаем к чтению аннотацию, описание, краткое содержание или предисловие (зависит от того, что написал сам автор книги «Перспективы отбора. От зеленых пеночек и бессмысленного усложнения до голых землекопов и мутирующего человечества»). Если вы не нашли необходимую информацию о книге — напишите в комментариях, мы постараемся отыскать её.
Перспективы отбора. От зеленых пеночек и бессмысленного усложнения до голых землекопов и мутирующего человечества — читать онлайн ознакомительный отрывок
Ниже представлен текст книги, разбитый по страницам. Система сохранения места последней прочитанной страницы, позволяет с удобством читать онлайн бесплатно книгу «Перспективы отбора. От зеленых пеночек и бессмысленного усложнения до голых землекопов и мутирующего человечества», без необходимости каждый раз заново искать на чём Вы остановились. Поставьте закладку, и сможете в любой момент перейти на страницу, на которой закончили чтение.
Интервал:
Закладка:
Эусоциальность— «высшая» форма социальности, при которой в социуме выделяется репродуктивная (размножающаяся) каста, а остальные особи отказываются от собственного размножения, посвящая свою жизнь заботе о семье. Эусоциальны, например, пчелы, муравьи, термиты, голые землекопы. В последние годы стало набирать популярность более широкое определение эусоциальности, под которое подпадает даже человек (у которого нет неразмножающейся касты, хотя иногда предлагают рассматривать в качестве таковой женщин после менопаузы). Мы в этой книге используем традиционное, узкое определение.
Эффективная численность популяции— сложное понятие из популяционной генетики, которое в самом первом приближении можно представить себе как численность особей, участвующих в размножении.
Вкладка
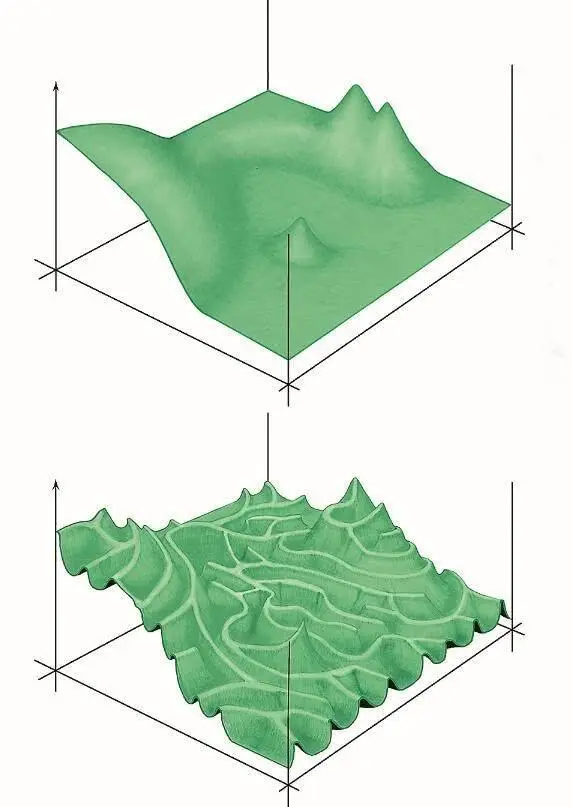
Илл. I.Примерно так можно изобразить ландшафт приспособленности.
Два горизонтальных измерения символизируют «пространство последовательностей», то есть все возможные варианты аминокислотной последовательности белка.
В действительности пространство последовательностей многомерно: в нем столько же измерений, сколько аминокислот в белке, и по каждому измерению белок может занимать одну из 20 позиций. Но такое пространство невозможно изобразить на бумаге, поэтому для простоты и наглядности его обычно схлопывают в два измерения.
Вертикальное измерение — эффективность выполнения белком своей функции (здесь мы опять сильно упрощаем реальность, предполагая, что чем эффективнее белок выполняет свою функцию, тем выше приспособленность организма). Естественный отбор двигает эволюционирующий белок вверх по склонам и препятствует спускам. Если эффекты мутаций слабо зависят друг от друга, ландшафт оказывается легкопроходимым ( график вверху ), что позволяет эволюции быстро освоить все потенциально доступные области.
В случае сильного эпистаза ландшафт превращается в лабиринт тропинок, разделенных пропастями ( график внизу ). В такой ситуации многие области пространства последовательностей могут остаться не разведанными эволюцией — просто потому, что до них трудно добраться.
Рисунок Андрея Томиленко.
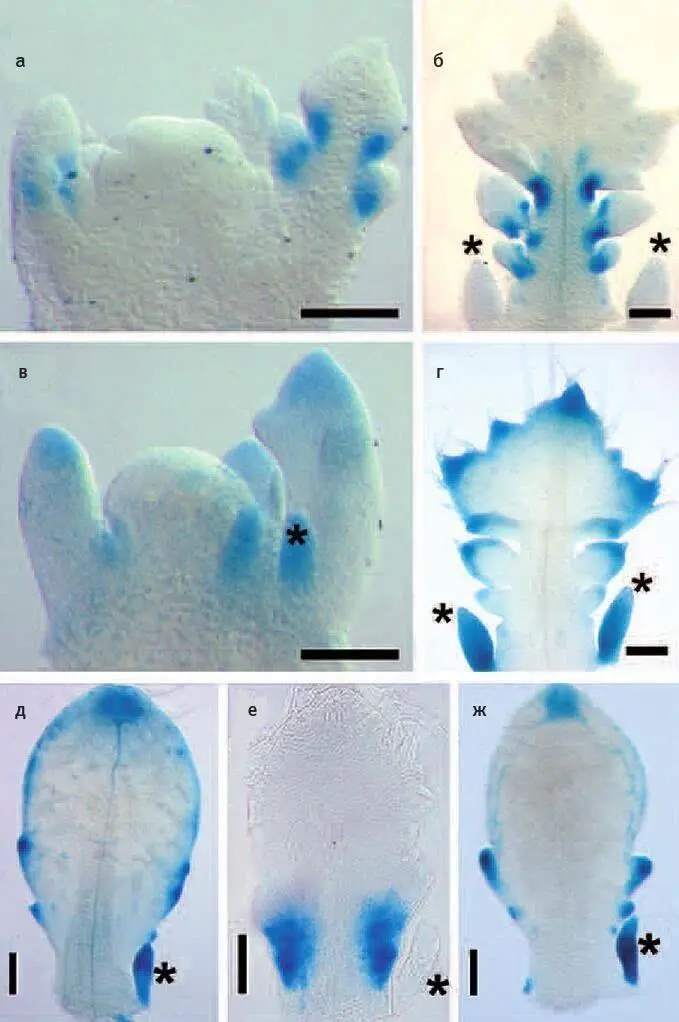
Илл. II.Области экспрессии генов RCO и LMI1. а — г : в зачатках листьев сердечника регуляторная область RCO обеспечивает экспрессию в основаниях лопастей (будущих вторичных листочков; а, б ), а регуляторная область LMI1 срабатывает на концах лопастей ( в, г ). д — ж : в зачатках листьев резуховидки регуляторная область LMI1 ( д — своя, ж — сердечника) обеспечивает экспрессию на конце листа и на вершинах мелких выступов по краю листовой пластинки, а регуляторная область RCO сердечника ( е ) срабатывает во внутренних областях листовой пластинки у ее основания. Звездочками отмечены прилистники. Длина масштабных линеек — 100 мкм. Из Vlad et al., 2014.

Илл. III.Полиморфизм окраски у тлей Macrosiphoniella yomogicola.

Илл. IV.Тринидадские гуппи — удобный объект для изучения быстрых эволюционных изменений. Из синопсиса к Ghalambor et al., 2015.
С сайта blog.tamagaro.net.

Илл. V.Восхитительные узоры на крыльях бабочек складываются из отдельных чешуек, различающихся по цвету, форме и строению. Окраска одних чешуек определяется пигментами, других — тонкослойной интерференцией. Каждая чешуйка одноцветна и образуется единственной клеткой.
Cornel Constantin / Используется по лицензии Shutterstock.com.

Илл. VI.Примеры параллельной эволюции цихлид в трех больших африканских озерах. Звездочками отмечены виды, геномы которых были прочтены.
По рисунку из синопсиса к Brawand et al., 2014.
Читать дальшеИнтервал:
Закладка:
Похожие книги на «Перспективы отбора. От зеленых пеночек и бессмысленного усложнения до голых землекопов и мутирующего человечества»
Представляем Вашему вниманию похожие книги на «Перспективы отбора. От зеленых пеночек и бессмысленного усложнения до голых землекопов и мутирующего человечества» списком для выбора. Мы отобрали схожую по названию и смыслу литературу в надежде предоставить читателям больше вариантов отыскать новые, интересные, ещё непрочитанные произведения.










Обсуждение, отзывы о книге «Перспективы отбора. От зеленых пеночек и бессмысленного усложнения до голых землекопов и мутирующего человечества» и просто собственные мнения читателей. Оставьте ваши комментарии, напишите, что Вы думаете о произведении, его смысле или главных героях. Укажите что конкретно понравилось, а что нет, и почему Вы так считаете.