Л. Наумова - Основы общей экологии
Здесь есть возможность читать онлайн «Л. Наумова - Основы общей экологии» весь текст электронной книги совершенно бесплатно (целиком полную версию без сокращений). В некоторых случаях можно слушать аудио, скачать через торрент в формате fb2 и присутствует краткое содержание. Город: Москва, Год выпуска: 2003, Издательство: Логос, Жанр: Современная проза, на русском языке. Описание произведения, (предисловие) а так же отзывы посетителей доступны на портале библиотеки ЛибКат.

- Название:Основы общей экологии
- Автор:
- Издательство:Логос
- Жанр:
- Год:2003
- Город:Москва
- ISBN:нет данных
- Рейтинг книги:3 / 5. Голосов: 1
-
Избранное:Добавить в избранное
- Отзывы:
-
Ваша оценка:
Основы общей экологии: краткое содержание, описание и аннотация
Предлагаем к чтению аннотацию, описание, краткое содержание или предисловие (зависит от того, что написал сам автор книги «Основы общей экологии»). Если вы не нашли необходимую информацию о книге — напишите в комментариях, мы постараемся отыскать её.
Для студентов высших учебных заведений, обучающихся по направлениям и специальностям: «Экология», «Биология», «Медицина», «Сельское хозяйство». Представляет интерес для научных работников в области биологии, экологии, лесного и сельского хозяйства.
Основы общей экологии — читать онлайн бесплатно полную книгу (весь текст) целиком
Ниже представлен текст книги, разбитый по страницам. Система сохранения места последней прочитанной страницы, позволяет с удобством читать онлайн бесплатно книгу «Основы общей экологии», без необходимости каждый раз заново искать на чём Вы остановились. Поставьте закладку, и сможете в любой момент перейти на страницу, на которой закончили чтение.
Интервал:
Закладка:
Такая циклическая динамика наблюдается, например, в популяциях тундровых леммингов, которые питаются мхами и лишайниками. Они продолжают активную жизнедеятельность под снегом и выедают свою кормовую базу настолько, что прекращают размножаться, а затем начинают умирать от бескормицы. После того, как мхи отрастут, начинается новый подъем численности леммингов.
Возможны колебания численности популяции под влиянием погодных условий, паразитов и хищников.
Существует особый вариант регулирования плотности популяций, который называется «оппортунистическим», т.е. не укладывающимся в «правильные» законы, описанные экспоненциальной или логистической кривой.
У эксплерентов (r-стратегов) происходят вспышки численности в том случае, если появляются обильные ресурсы. При этом рост численности происходит либо за счет того, что начинают развиваться особи из покоящихся диаспор (скажем, почвенного банка семян), либо за счет массового «десанта» в стадии яиц (скажем, мух, налетевших на труп животного). Так как конкуренция в силу обилия ресурсов слаба то израсходовав их изобилие, популяция погибает целиком.
У растений-эксплерентов при повышении плотности популяции конкуренция возрастает, но самоизреживания (как у деревьев-виолентов) не происходит, а уменьшается размер особей в десятки и сотни раз. При этом растения проходят весь жизненный цикл и способны дать семена.
Дж. Харпер (Harper, 1977) назвал такой тип регуляции плотности популяций растений-однолетников «пластичностью» и противопоставил его самоизреживанию. Эти два типа регулирования плотности в популяциях растений связаны переходом: у большинства видов с вторичными стратегиями при повышении плотности популяции происходит одновременно и уменьшение размера особей, и самоизреживание.
На знании этих закономерностей построено обоснование нормы высева культурных растений. Вначале при увеличении нормы высева урожай растет, но потом начинает снижаться (рис. 17). Выбирается та норма высева, которая обеспечивает максимальный урожай. Впрочем, иногда ее несколько завышают, чтобы культурные растения могли подавлять популяции сорных растений. При усилении гербицидного контроля в этом нет необходимости.
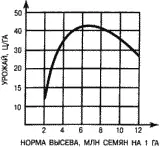
Рис. 17. Зависимость урожая пшеницы от нормы высева при оптимальных экологических условиях.
1. Охарактеризуйте экспоненциальную модель роста популяции.
2. Почему модель экспоненциального роста редко наблюдается в природных популяциях?
3. Из каких фаз состоит логистическая модель роста популяции?
4. Какие причины вызывают циклическую динамику популяций?
5. Какие популяции называются оппортунистическими?
7.4. Возрастной состав популяций
Кривые выживания могут реализовываться при разном характере динамики популяций: при одновременном «старте» популяций, заселяющих свободное пространство, или при постоянном «популяционном потоке», когда часть особей вымирает, а часть – занимает освободившееся место (ситуация «вокзала», в которой число уезжающих пассажиров постоянно компенсируется вновь прибывающими). В результате в разных популяциях при единовременном учете выявляется разный возрастной состав.
Определить абсолютный возраст особи можно не у всех видов. Это несложно сделать для деревьев, воспользовавшись специальным буром, которым извлекают столбик древесины – керн и подсчитывают на нем число годичных колец. У дерева конкретного вида в конкретных условиях (т.е. при одном бонитете) можно определить возраст с точностью до 5 лет по диаметру ствола. У хвойных возраст определяется по числу мутовок побегов на стволе. Однако у трав определение абсолютного возраста затруднительно, и потому оценивают их «возрастное состояние» (стадию).
У высших споровых растений (папоротники, хвощи, плауны) выделяются стадии спор, заростков-гаметофитов, молодых спорофитов и взрослых спорофитов.
Большой опыт выделения возрастных состояний растений накоплен ботаниками-демографами (Л.Б. Заугольновой, О.В. Смирновой, Л.И. Жуковой и др., табл. 7).
Таблица 7 Периодизация онтогенеза цветковых растений (по Жуковой, 1987)
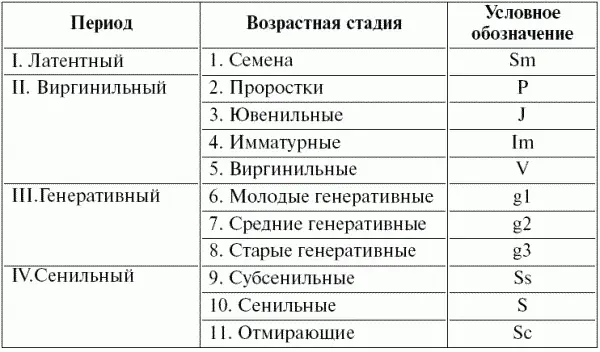
По информативности оценка «возрастного состояния» во многом уступает оценке абсолютного возраста, так как в плохих условиях молодое растение может «выглядеть старым», и, наоборот, при хороших условиях за растение среднего возраста можно принять молодую особь. Соответственно возможны ошибки и при определении возрастного состава популяций и воценке тенденций их динамики.
Читать дальшеИнтервал:
Закладка:
Похожие книги на «Основы общей экологии»
Представляем Вашему вниманию похожие книги на «Основы общей экологии» списком для выбора. Мы отобрали схожую по названию и смыслу литературу в надежде предоставить читателям больше вариантов отыскать новые, интересные, ещё непрочитанные произведения.












Обсуждение, отзывы о книге «Основы общей экологии» и просто собственные мнения читателей. Оставьте ваши комментарии, напишите, что Вы думаете о произведении, его смысле или главных героях. Укажите что конкретно понравилось, а что нет, и почему Вы так считаете.