Moselio Schaechter - In the Company of Microbes
Здесь есть возможность читать онлайн «Moselio Schaechter - In the Company of Microbes» — ознакомительный отрывок электронной книги совершенно бесплатно, а после прочтения отрывка купить полную версию. В некоторых случаях можно слушать аудио, скачать через торрент в формате fb2 и присутствует краткое содержание. Жанр: unrecognised, на английском языке. Описание произведения, (предисловие) а так же отзывы посетителей доступны на портале библиотеки ЛибКат.

- Название:In the Company of Microbes
- Автор:
- Жанр:
- Год:неизвестен
- ISBN:нет данных
- Рейтинг книги:3 / 5. Голосов: 1
-
Избранное:Добавить в избранное
- Отзывы:
-
Ваша оценка:
In the Company of Microbes: краткое содержание, описание и аннотация
Предлагаем к чтению аннотацию, описание, краткое содержание или предисловие (зависит от того, что написал сам автор книги «In the Company of Microbes»). Если вы не нашли необходимую информацию о книге — напишите в комментариях, мы постараемся отыскать её.
In the Company of Microbes Join Schaechter in discovering a never-ending pageant of astounding variations on the theme of microbial life. Enjoy!
In the Company of Microbes — читать онлайн ознакомительный отрывок
Ниже представлен текст книги, разбитый по страницам. Система сохранения места последней прочитанной страницы, позволяет с удобством читать онлайн бесплатно книгу «In the Company of Microbes», без необходимости каждый раз заново искать на чём Вы остановились. Поставьте закладку, и сможете в любой момент перейти на страницу, на которой закончили чтение.
Интервал:
Закладка:
Domestication has led to the production of long chains of sessile cells. Shown is a fluorescence micrograph taken by Dan Kearns of growing cells of laboratory B. subtilis . In addition to swimming cells (the green-colored singlets and doublets), the population contains many long chains of sessile cells. The cells were visualized with the vital membrane stain FM4-64 (red) and contained a fusion of the gene for the Green Fluorescence Protein (responsible for the green color) to a promoter under the control of a transcription factor that controls motility. Thus, only the motile cells in the image are green. Wild (undomesticated) strains, in comparison, produce relatively few chains of sessile cells.
Ferdinand Cohn reported the discovery of B. subtilis in 1877. But the B. subtilis laboratory strains of today are a shadow of their former selves. Years and years of manipulation in the laboratory has robbed B. subtilis of much of its biology. On the one hand, laboratory strains can be transformed with DNA much more efficiently than undomesticated strains. On the other hand, laboratory strains are generally deficient in a variety of behaviors manifest in wild strains. Whereas wild strains are highly motile, have the capacity to swarm on surfaces, and form architecturally complex communities (biofilms), laboratory strains form long chains of sessile cells, fail to swarm, and form smooth colonies and thin pellicles. Indeed, studies with biofilms (in collaboration with Roberto Kolter) have changed our thinking about sporulation. We traditionally treated sporulation as largely a behavior of solitary cells, but recent work emphasizes the importance of studying spore formation in the context of multicellular communities (as has long been recognized in myxobacteria).
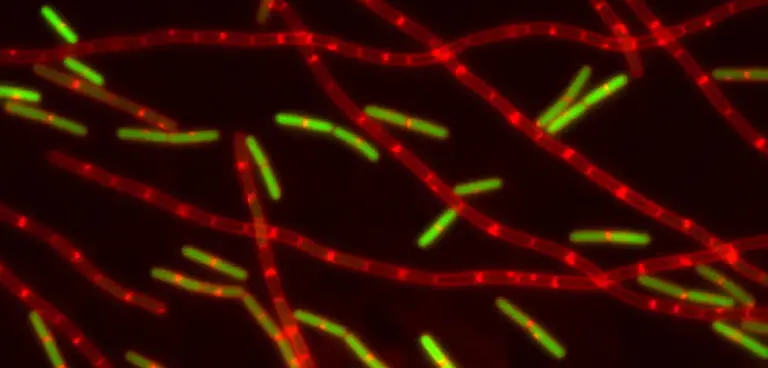
Domestication has led to the production of long chains of sessile cells. Shown is a fluorescence micrograph taken by Dan Kearns of growing cells of laboratory B. subtilis. In addition to swimming cells (the green-colored singlets and doublets), the population contains many long chains of sessile cells. The cells were visualized with the vital membrane stain FM4-64 (red) and contained a fusion of the gene for the Green Fluorescence Protein (responsible for the green color) to a promoter under the control of a transcription factor that controls motility. Thus, only the motile cells in the image are green. Wild (undomesticated) strains, in comparison, produce relatively few chains of sessile cells.
Source: Kearns, DB, R Losick.2005. Cell population heterogeneity during growth of Bacillus subtilis. Genes & Development 19:3083-3094.
How did this happen? I suspect that it is the result of heavy mutagenesis along with generations of researchers who, without thinking about it, favored rare smooth colonies that stayed put on the agar plate over other more unruly ones.
There are two lessons here that may be of general interest for those of us who consider small things. First, much biology may await discovery from revisiting the ancestral roots of popular laboratory strains. Second, this missing biology may hold the key to understanding the function of some of the myriad mysterious genes with which bacterial genomes are riddled. In short, back to the wild!

Richard Losick is Harvard College Professor, Maria Moors Cabot Professor of Biology at Harvard University.
September 15, 2008
bit.ly/1M2m15Q
#8

by Elio
Can you think of a place on Earth where there is free water but no microbes? (outside the bodies of other organisms or the lab)
March 2, 2007
bit.ly/1Gfpsdq
7
The Tyranny of Phylogeny: An Exhortation
by Elio
Two archaea walk into a bar and the bartender says, “If you guys are going to start in with the jokes again, Woese is me.”
—Fred Rosenberg
There are days when I wish that the Woesian Three Domain scheme were wrong. Not that I would be happier if there were four or five or whatever number of domains. What would please me would be an escape from what I feel is an unnecessarily oppressive way of thinking, the seating of phylogeny (and its acolyte, genomics) alone at the head of the table. Why do I say this? Because as essential as phylogeny is to our understanding of the evolution of living organisms, equally vital is ecology to comprehend present day life. While it’s good to know where you come from, it’s equally important to know where you are and what you’re doing there. The Spanish philosopher Ortega y Gasset said it well: “ Yo soy yo y mi circunstancia ” (“ I am I and my circumstance ”).
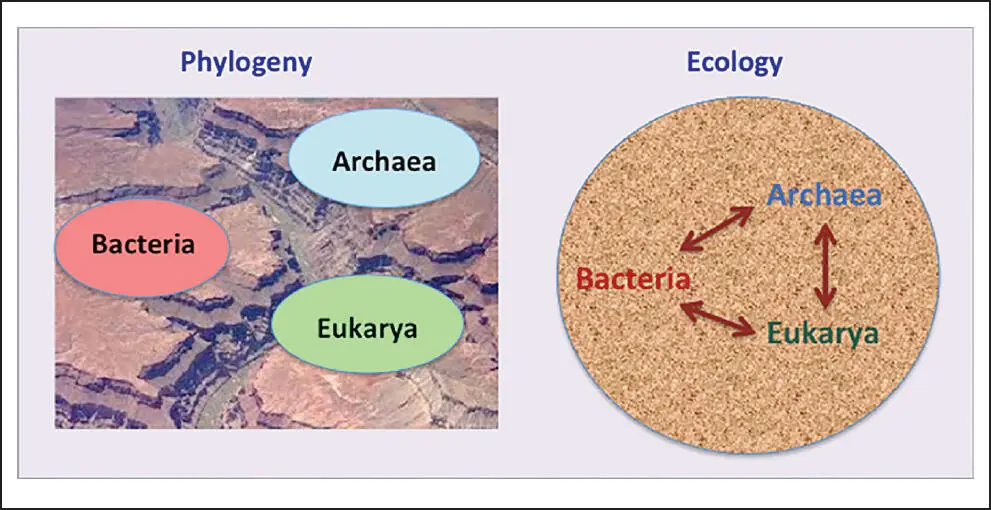
A sole focus on phylogeny forces the past ahead of the present. Compare the two figures; they represent two different worldviews. One highlights the deep clefts between the three domains, the other is integrative and does away with such barriers. (The ecology perspective also acknowledges the highly abundant viruses, indicated by the stippled background.) Maybe it is not phylogeny’s job to emphasize ecology, but neither should we be fixated on evolution alone. Obviously, no one is, so I apologize for erecting a straw man. Yet let me voice a wish: I would like to see a wider-ranging acknowledgement of each organism’s give-and-take with its environment. Famed evolutionary biologist and geneticist Dobzhansky once said: “ Nothing in Biology makes sense except in the light of evolution. ” Do I dare modify this well-known dictum to read: “ Nothing in Biology makes sense except in the light of evolution and ecology .”
October 25, 2012
bit.ly/1XgDLTc
8
Virus in the Room
by Welkin Johnson
I am the Lorax, I speak for the trees. I speak for the trees, for the trees have no tongues, and I’m asking you, sir, at the top of my lungs.
– from The Lorax by Dr. Seuss
As biologists, we divvy the biological realm up into domains using a formula that, frankly, smacks of nepotism, bestowing three glorious domains upon our closest relatives—the Eucaryota, the Archaea, and the Bacteria—while committing an injustice to the so-called viruses, lumping them together in a miscellaneous catch-all category (“viruses” from Latin for poison and other noxious substances) with contemptible disregard for phylogeny or any true measure of diversity.
Imagine that viruses, like Dr. Seuss’s truffula trees, had a vocal advocate like the Lorax. Undoubtedly, through the agency of their outspoken mouthpiece, they would protest these gerrymandered borders and laugh at our skewed notions of biological diversity. After all (the viruses would argue), just consider the platypus, the coelacanth, the earthworm, and the bacillus. All these organisms have double-stranded DNA genomes, whose lengths all fall within roughly the same order of magnitude, which they replicate using evolutionarily customized versions of what amounts to the same basic enzymatic apparatus. How boring! How unimaginative! Now consider this (the viruses go on to say): the giant Mimivirus, 1256 nm of girth enfolding >1,000,000 base pairs of DNA, and the tiny Circovirus, with a mere 1,800 bases of single-stranded DNA tucked inside a 20 nm-wide shell, are neither more nor less related to one another than either one is to an elephant! (For those who are not familiar with the elephant, it is a relative of the platypus, the coelacanth, the earthworm, and the bacillus.)
Читать дальшеИнтервал:
Закладка:
Похожие книги на «In the Company of Microbes»
Представляем Вашему вниманию похожие книги на «In the Company of Microbes» списком для выбора. Мы отобрали схожую по названию и смыслу литературу в надежде предоставить читателям больше вариантов отыскать новые, интересные, ещё непрочитанные произведения.












Обсуждение, отзывы о книге «In the Company of Microbes» и просто собственные мнения читателей. Оставьте ваши комментарии, напишите, что Вы думаете о произведении, его смысле или главных героях. Укажите что конкретно понравилось, а что нет, и почему Вы так считаете.