Tina M. Henkin - Snyder and Champness Molecular Genetics of Bacteria
Здесь есть возможность читать онлайн «Tina M. Henkin - Snyder and Champness Molecular Genetics of Bacteria» — ознакомительный отрывок электронной книги совершенно бесплатно, а после прочтения отрывка купить полную версию. В некоторых случаях можно слушать аудио, скачать через торрент в формате fb2 и присутствует краткое содержание. Жанр: unrecognised, на английском языке. Описание произведения, (предисловие) а так же отзывы посетителей доступны на портале библиотеки ЛибКат.

- Название:Snyder and Champness Molecular Genetics of Bacteria
- Автор:
- Жанр:
- Год:неизвестен
- ISBN:нет данных
- Рейтинг книги:3 / 5. Голосов: 1
-
Избранное:Добавить в избранное
- Отзывы:
-
Ваша оценка:
Snyder and Champness Molecular Genetics of Bacteria: краткое содержание, описание и аннотация
Предлагаем к чтению аннотацию, описание, краткое содержание или предисловие (зависит от того, что написал сам автор книги «Snyder and Champness Molecular Genetics of Bacteria»). Если вы не нашли необходимую информацию о книге — напишите в комментариях, мы постараемся отыскать её.
Although the text is centered on the most-studied bacteria,
and
, many examples are drawn from other bacteria of experimental, medical, ecological, and biotechnological importance. The book's many useful features include
Text boxes to help students make connections to relevant topics related to other organisms, including humans A summary of main points at the end of each chapter Questions for discussion and independent thought A list of suggested readings for background and further investigation in each chapter Fully illustrated with detailed diagrams and photos in full color A glossary of terms highlighted in the text While intended as an undergraduate or beginning graduate textbook, Molecular Genetics of Bacteria is an invaluable reference for anyone working in the fields of microbiology, genetics, biochemistry, bioengineering, medicine, molecular biology, and biotechnology.
"This is a marvelous textbook that is completely up-to-date and comprehensive, but not overwhelming. The clear prose and excellent figures make it ideal for use in teaching bacterial molecular genetics."—
, University of Washington
Snyder and Champness Molecular Genetics of Bacteria — читать онлайн ознакомительный отрывок
Ниже представлен текст книги, разбитый по страницам. Система сохранения места последней прочитанной страницы, позволяет с удобством читать онлайн бесплатно книгу «Snyder and Champness Molecular Genetics of Bacteria», без необходимости каждый раз заново искать на чём Вы остановились. Поставьте закладку, и сможете в любой момент перейти на страницу, на которой закончили чтение.
Интервал:
Закладка:
Figure 1.23shows the results of these experiments. For convenience, the following letters were assigned to each of the intervals during the cell cycle. The letter I denotes the time from when the last round of chromosome replication initiated until a new round begins. C is the time it takes to replicate the entire chromosome, and D is the time from when a round of chromosome replication is completed until cell division occurs. The top of the figure shows the relationship of I, C, and D when cells are growing slowly, with a generation time of 70 min. Under these conditions, I is 70 min, C is 40 min, and D is 20 min. However, when the cells are growing in a richer medium and are dividing more rapidly, with a generation time of only 30 min, the pattern changes. The C and D intervals remain about the same, but the I interval is much shorter, only about 30 min.
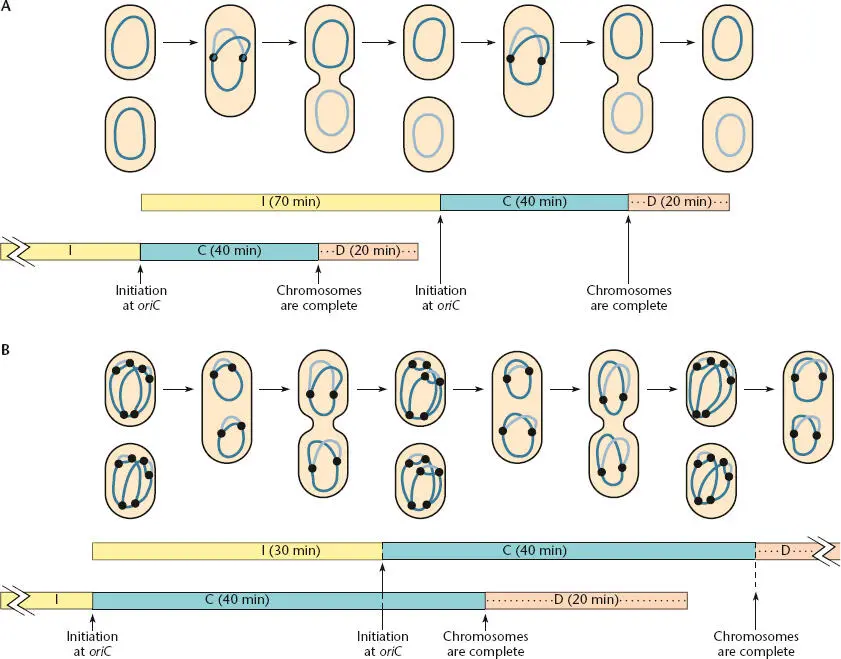
Figure 1.23 Timing of DNA replication during the cell cycle, with two different generation times (A and B). The time between initiations (I) is the only time that changes. See the text for definitions of I, C, and D.
Some conclusions may be drawn from these data. One conclusion is that the C and D intervals remain about the same independent of the growth rate. At 37°C, the time it takes the chromosome to replicate is always about 40 min, and it takes about 20 min from the time a round of replication terminates until the cell divides. However, the I interval gets shorter when the cells are growing faster and have shorter generation times. In fact, the I interval is approximately equal to the generation time—the time it takes a newborn cell to grow and divide. This makes sense because, as discussed below, initiation of chromosome replication appears to occur every time the cells reach a certain size. They reach this size once every generation time, independent of how fast they are growing.
Another point apparent from the data is that in cells growing rapidly with a short generation time, the I interval can be shorter than the C interval. If I is shorter than C, a new round of chromosomal DNA replication will begin before the old one is completed. This explains the higher DNA content of fast-growing cells as compared to slowgrowing cells. It also explains the observation that genes closer to the origin of replication are present in more copies than are genes closer to the replication terminus.
Despite providing these important results, this elegant analysis does not allow us to tell whether division is coupled to initiation or termination of chromosomal DNA replication. The fact that the I interval always equals the generation time suggests that the events leading up to division are set in motion at the time a round of chromosome replication is initiated and are completed 60 min later independent of how fast the cells are growing. However, it is also possible that the act of termination of a round of chromosome replication sets in motion a cell division 20 min later. Multiple laboratories are working to resolve these issues.
Timing of Initiation of Replication
A new round of replication must be initiated each time the cell divides, or the amount of DNA in the cell would increase until the cells were stuffed full of it or decrease until no cell had a complete copy of the chromosome. Clearly, initiation of replication is exquisitely timed. In cells growing very rapidly, in which the next rounds of replication initiate before the last ones are completed, so that the cells contain a number of origins of replication, all of the origins in a cell “fire” simultaneously, indicating tight control.
A number of attempts have been made to correlate the timing of initiation of chromosome replication with other cellular parameters during the cell cycle. Most evidence from such attempts points to initiation of replication being tied to cell mass. After cells divide, their mass, or weight, continuously increases until they divide again. The initiation of chromosome replication occurs each time the cell achieves a certain mass, the initiation mass. If cells are growing faster in richer medium, they are larger and achieve the initiation mass sooner than do smaller, slower-growing cells, explaining why new rounds of chromosome replication occur before the termination of previous rounds in faster-growing cells but not in slower-growing cells. However, these experiments by themselves do not explain what it is about the cell mass that triggers initiation.
ROLE OF THE DnaA PROTEIN
DnaA is essential for initiating DNA replication at oriC and along with other components also helps to regulate the timing of DNA replication. DnaA primarily affects the frequency of initiation by changes in the ATP-binding versus ADP-binding state of DnaA and by access of DnaA to oriC . DnaA must be in its ATP-bound state to form the structure used to separate the DNA strands at oriC to initiate DNA replication. DnaA hydrolyzes ATP slowly; therefore, other factors are required to stimulate its ATPase activity to inactivate it until it is needed for the next round of DNA replication. The affinity of DnaA for the various binding sites within oriC (see “Origin of Chromosomal Replication” above) allows a mechanism for the origin to be identified but not immediately used as an origin until all conditions are in place for replication. A number of processes control the pool of DnaA found in the ATP-bound state. Reducing the pool of DnaA-ATP following initiation is an important mechanism for delaying reinitiation, thereby allowing replication to proceed in an orderly fashion. The production of new DnaA also plays a role in the ratio of ATP-bound versus ADP-bound DnaA, and because of the high ATP concentration in cells, newly made DnaA will be bound by ATP.
Inactivation of DnaA by hydrolysis to DnaA-ADP
Conversion to the DnaA-ADP state occurs primarily by two mechanisms following initiation of DNA replication. As mentioned above, only the ATP-bound form can bind all of the DnaA boxes in oriC . Only when all of the sites are bound can the structure form that will be needed to open the DUE to allow replication to initiate ( Figure 1.15). This helps the DnaA protein to act as a switch that is largely independent of its concentration because other cellular inputs can control the ATP-bound versus ADP-bound state of DnaA. One cellular input that reduces the pool in the DnaA-ATP state involves the presence of the assembled replication fork at the origin region immediately after initiation of replication. The presence of the β sliding-clamp protein causes DnaA to hydrolyze ATP to ADP by interacting with another protein, a relative of DnaA called Hda (for homology to DnaA) (see Camara et al., Suggested Reading). This process is sometimes referred to as regulatory inactivation of DnaA, or RIDA. A second input that reduces the pool of DnaA-ATP involves a locus in the chromosome called datA . The datA locus, which is about 1 kb in length, has five DnaA-binding sites and a binding site for IHF. Binding of DnaA and IHF to this locus induces an endogenous ATPase activity of the DnaA protein, coaxing it into the inactive DnaA-ADP form in a process abbreviated as DDAH ( datA -dependent DnaA-ATP hydrolysis). How specifically DDAH is controlled with the cell cycle remains unknown.
Reactivation of DnaA by nucleotide exchange to DnaA-ATP
Production of new DnaA in the ATP-bound form is one mechanism for increasing the DnaA-ATP/DnaA-ADP ratio in cells. There are also two mechanisms involved in “recycling” DnaA by encouraging it to exchange a nucleotide, putting it back into the DnaA-ATP form; one involves an exchange catalyzed by acidic phospholipids, and the other involves two sites found in the chromosome called DnaA-reactivating sequences (see Fujimitsu et al., Suggested Reading). The numerous inputs work together to limit the initiation of DNA replication to help keep the number of chromosomes consistent with the requirements for cell division.
Читать дальшеИнтервал:
Закладка:
Похожие книги на «Snyder and Champness Molecular Genetics of Bacteria»
Представляем Вашему вниманию похожие книги на «Snyder and Champness Molecular Genetics of Bacteria» списком для выбора. Мы отобрали схожую по названию и смыслу литературу в надежде предоставить читателям больше вариантов отыскать новые, интересные, ещё непрочитанные произведения.











Обсуждение, отзывы о книге «Snyder and Champness Molecular Genetics of Bacteria» и просто собственные мнения читателей. Оставьте ваши комментарии, напишите, что Вы думаете о произведении, его смысле или главных героях. Укажите что конкретно понравилось, а что нет, и почему Вы так считаете.