Jane Flint - Principles of Virology, Volume 1
Здесь есть возможность читать онлайн «Jane Flint - Principles of Virology, Volume 1» — ознакомительный отрывок электронной книги совершенно бесплатно, а после прочтения отрывка купить полную версию. В некоторых случаях можно слушать аудио, скачать через торрент в формате fb2 и присутствует краткое содержание. Жанр: unrecognised, на английском языке. Описание произведения, (предисловие) а так же отзывы посетителей доступны на портале библиотеки ЛибКат.

- Название:Principles of Virology, Volume 1
- Автор:
- Жанр:
- Год:неизвестен
- ISBN:нет данных
- Рейтинг книги:5 / 5. Голосов: 1
-
Избранное:Добавить в избранное
- Отзывы:
-
Ваша оценка:
Principles of Virology, Volume 1: краткое содержание, описание и аннотация
Предлагаем к чтению аннотацию, описание, краткое содержание или предисловие (зависит от того, что написал сам автор книги «Principles of Virology, Volume 1»). Если вы не нашли необходимую информацию о книге — напишите в комментариях, мы постараемся отыскать её.
Volume I: Molecular Biology
Volume II: Pathogenesis and Control
Principles of Virology, Fifth Edition
Principles of Virology, Volume 1 — читать онлайн ознакомительный отрывок
Ниже представлен текст книги, разбитый по страницам. Система сохранения места последней прочитанной страницы, позволяет с удобством читать онлайн бесплатно книгу «Principles of Virology, Volume 1», без необходимости каждый раз заново искать на чём Вы остановились. Поставьте закладку, и сможете в любой момент перейти на страницу, на которой закончили чтение.
Интервал:
Закладка:
The genome structure of this virus suggests that at some point in the past a flavivirus genome fragmented to produce the RNA segments encoding the NS3and NS5-like proteins. This fragmentation might have initially taken place as shown for foot-and-mouth disease virus in cells in culture, by fixing of deletion mutations that complemented one another. Next, coinfection of this segmented flavivirus with another unidentified virus could have produced the precursor of Jingmen tick virus.
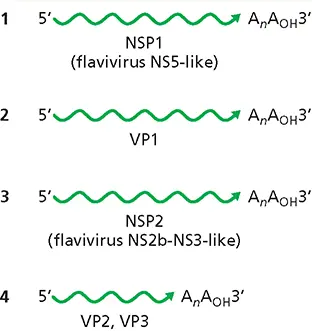
RNA genome of JMTV virus.The viral genome comprises four segments of single-stranded, (+) sense RNA. Proteins encoded by each RNA are indicated. RNA segments 1 and 3 encode flavivirus-like proteins.
The results provide new clues about the origins of segmented RNA viruses.
Moreno E, Ojosnegros S, García-Arriaza J, Escarmís C, Domingo E, Perales C. 2014. Exploration of sequence space as the basis of viral RNA genome segmentation. Proc Natl Acad Sci U S A 111:6678–6683.
Qin XC, Shi M, Tian JH, Lin XD, Gao DY, He JR, Wang JB, Li CX, Kang YJ, Yu B, Zhou DJ, Xu J, Plyusnin A, Holmes EC, Zhang YZ. 2014. A tick-borne segmented RNA virus contains genome segments derived from unsegmented viral ancestors. Proc Natl Acad Sci U S A 111:6744–6749.
Classical Genetic Methods
Mapping Mutations
Before the advent of recombinant DNA technology, it was extremely difficult for investigators to determine the locations of mutations in viral genomes. The marker rescuetechnique (described in “Introducing Mutations into the Viral Genome” below) was a solution to this problem, but before it was developed, other, less satisfactory approaches were exploited.
Recombination mapping can be applied to both DNA and RNA viruses. Recombination results in genetic exchange between genomes within the infected cell. The frequency of recombination between two mutations in a linear genome increases with the physical distance separating them. In practice, cells are coinfected with two mutants, and the frequency of recombination is calculated by dividing the titer of phenotypically wild-type virus ( Box 3.7) obtained under restrictive conditions (e.g., high temperature) by the titer measured under permissive conditions (e.g., low temperature). The recombination frequency between pairs of mutants is determined, allowing the mutations to be placed on a contiguous map. Although a location can be assigned for each mutation relative to others, this approach does not result in a physical map of the actual location of the base change in the genome.
In the case of RNA viruses with segmented genomes, the technique of reassortmentallows the assignment of mutations to specific genome segments. When cells are coinfected with both mutant and wild-type viruses, the progeny includes reassortantsthat inherit RNA segments from either parent. The origins of the RNA segments can be deduced from their migration patterns during gel electrophoresis ( Fig. 3.11) or by nucleic acid hybridization. By analyzing a panel of such reassortants, the segment responsible for the phenotype can be identified.
Functional Analysis
Complementationdescribes the ability of gene products from two different mutant viruses to interact functionally in the same cell, permitting viral reproduction. It can be distinguished from recombination or reassortment by examining the progeny produced by coinfected cells. True complementation yields only the two parental mutants, while wild-type genomes result from recombination or reassortment. If the mutations being tested are in separate genes, each virus is able to supply a functional gene product, allowing both viruses to be reproduced. If the two viruses carry mutations in the same gene, no reproduction will occur. In this way, the members of collections of mutants obtained by chemical mutagenesis were initially organized into complementation groups defining separate viral functions. In principle, there can be as many complementation groups as genes.
BOX 3.6
METHODS
Spontaneous and induced mutations
In the early days of experimental virology, mutant viruses could be isolated only by screening stocks for interesting phenotypes, for none of the tools that we now take for granted, such as restriction endonucleases, efficient DNA sequencing methods, and molecular cloning procedures, were developed until the mid to late 1970s. RNA virus stocks usually contain a high proportion of mutants, and it is only a matter of devising the appropriate selection conditions (e.g., high or low temperature or exposure to drugs that inhibit viral reproduction) to select mutants with the desired phenotype from the total population. For example, the live attenuated poliovirus vaccine strains developed by Albert Sabin are mutants that were selected from a virulent virus stock (Volume II, Fig. 7.11).
The low spontaneous mutation rate of DNA viruses necessitated random mutagenesis by exposure to a chemical mutagen. Mutagens such as nitrous acid, hydroxylamine, and alkylating agents chemically modify the nucleic acid in preparations of virus particles, resulting in changes in base-pairing during subsequent genome replication. Base analogs, intercalating agents, or UV light are applied to the infected cell to cause changes in the viral genome during replication. Such agents introduce mutations more or less at random. Some mutations are lethal under all conditions, while others have no effect and are said to be silent.
To facilitate identification of mutants, the population must be screened for a phenotype that can be identified easily in a plaque assay. One such phenotype is temperature-sensitive viability of the virus. Virus mutants with this phenotype reproduce well at low temperatures, but poorly or not at all at high temperatures. The permissive and nonpermissive temperatures are typically 33 and 39°C, respectively, for viruses that replicate in mammalian cells. Other commonly sought phenotypes are changes in plaque size or morphology, drug resistance, antibody resistance, and host range (that is, loss of the ability to reproduce in certain hosts or host cells).
BOX 3.7
TERMINOLOGY
What is wild type?
Terminology can be confusing. Virologists often use terms such as “strains,” “variants,” and “mutants” to designate a virus that differs in some heritable way from a parental or wild-type virus. In conventional usage, the wild typeis defined as the original (often laboratory-adapted) virus from which mutants are selected and which is used as the basis for comparison. A wild-type virus may notbe identical to a virus isolated from nature. In fact, the genome of a wild-type virus may include numerous mutations accumulated during propagation in the laboratory. For example, the genome of the first isolate of poliovirus obtained in 1909 undoubtedly is very different from that of the virus we call wild type today. We distinguish carefully between laboratory wild types and new virus isolates from the natural host. The latter are called field isolatesor clinical isolates.
The field of viral taxonomy has its own naming conventions which can cause some confusion. Viruses are classified into orders, families, subfamilies, genera, and species. These names are always italicized and start with a capital letter (e.g., Picornaviridae). To ensure clarity, the names of viruses (like poliovirus) should be written differently from the names of species (which are constructs that assist in the cataloging of viruses). A species name is written in italics with the first word beginning with a capital letter (other words should be capitalized if they are proper nouns). For example, the causative agents of poliomyelitis, poliovirus types 1, 2, and 3, are members of the species Enterovirus C . A virus name should never be italicized, even when it includes the name of a host species or genus, and should be written in lowercase: for example, Sida ciliaris golden mosaic virus. A good exercise would be to see how often we have accidentally violated these rules in this textbook.
Читать дальшеИнтервал:
Закладка:
Похожие книги на «Principles of Virology, Volume 1»
Представляем Вашему вниманию похожие книги на «Principles of Virology, Volume 1» списком для выбора. Мы отобрали схожую по названию и смыслу литературу в надежде предоставить читателям больше вариантов отыскать новые, интересные, ещё непрочитанные произведения.



Обсуждение, отзывы о книге «Principles of Virology, Volume 1» и просто собственные мнения читателей. Оставьте ваши комментарии, напишите, что Вы думаете о произведении, его смысле или главных героях. Укажите что конкретно понравилось, а что нет, и почему Вы так считаете.