Коллектив авторов - Строение и история развития литосферы
Здесь есть возможность читать онлайн «Коллектив авторов - Строение и история развития литосферы» — ознакомительный отрывок электронной книги совершенно бесплатно, а после прочтения отрывка купить полную версию. В некоторых случаях можно слушать аудио, скачать через торрент в формате fb2 и присутствует краткое содержание. Город: Москва, Год выпуска: 2010, ISBN: 2010, Издательство: Array Литагент «Паулсен», Жанр: geography_book, на русском языке. Описание произведения, (предисловие) а так же отзывы посетителей доступны на портале библиотеки ЛибКат.

- Название:Строение и история развития литосферы
- Автор:
- Издательство:Array Литагент «Паулсен»
- Жанр:
- Год:2010
- Город:Москва
- ISBN:978-5-98797-043-0
- Рейтинг книги:4 / 5. Голосов: 1
-
Избранное:Добавить в избранное
- Отзывы:
-
Ваша оценка:
Строение и история развития литосферы: краткое содержание, описание и аннотация
Предлагаем к чтению аннотацию, описание, краткое содержание или предисловие (зависит от того, что написал сам автор книги «Строение и история развития литосферы»). Если вы не нашли необходимую информацию о книге — напишите в комментариях, мы постараемся отыскать её.
Строение и история развития литосферы — читать онлайн ознакомительный отрывок
Ниже представлен текст книги, разбитый по страницам. Система сохранения места последней прочитанной страницы, позволяет с удобством читать онлайн бесплатно книгу «Строение и история развития литосферы», без необходимости каждый раз заново искать на чём Вы остановились. Поставьте закладку, и сможете в любой момент перейти на страницу, на которой закончили чтение.
Интервал:
Закладка:
Микропалеонтологический анализ показал, что почти все образцы содержат планктонные и бентосные фораминиферы, кроме того в ряде горизонтов найдены диатомовые водоросли и наннопланктон. При этом количество микрофауны подвержено резким ритмическим колебаниям по разрезу (рис. 1). Выделяются 5 пиков обилия микропланктона, которые мы называем интервалами высокой продуктивности (ИВП = high productivity intervals, HP). В соответствующих горизонтах количество планктонных фораминифер составляет более 3 тыс. экз./1 г. Кроме этого в разрезе колонки фиксируются 6 горизонтов крайне обедненных микрофауной, формирование которых, по-видимому, происходило во время существования постоянного морского льда. Большинство из них узкие по мощности и два (35–110 см и 150–200 см) довольно мощные.
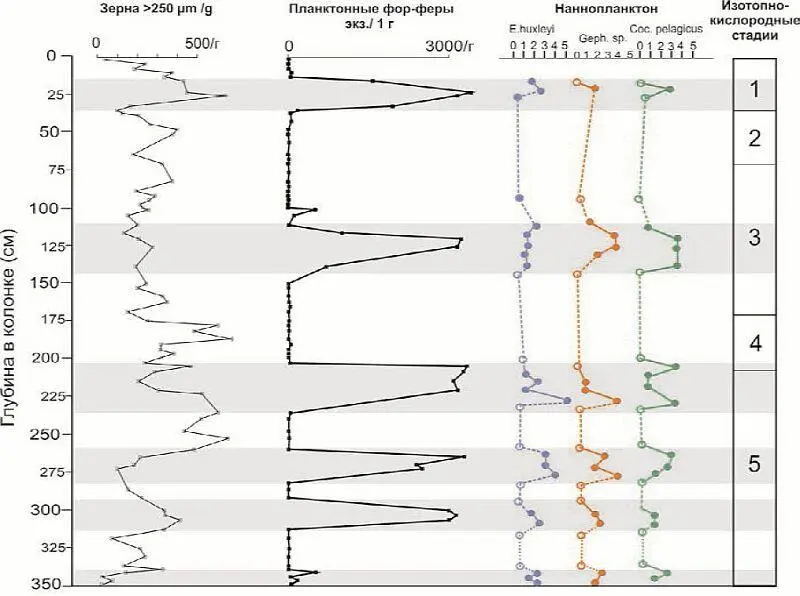
Рис. 1. Материал ледового разноса, планктонные фораминиферы и наннопланктон в колонке 2417 (хребет Книповича). Серым цветом выделены интервалы высокой продуктивности (ИВП).
Основную массу фораминиферового комплекса составляет планктонная арктическая левозавитая Neogloboquadrina pachyderma sin. Ее количество колеблется от 1–2 раковин до тысяч экземпляров в образце. В ИВП в комплексе, помимо обильных N. pachyderma sin, присутствуют единичные раковины более тепловодных N. pachyderma dex, Globigerina quinqueloba, G. bulloides (ювенильные), Globigerinita glutinata .
Три из этих интервалов выделяются присутствием особенно тепловодных видов планктонных фораминифер. В нижнем горизонте (337–350 см) это субтропические Globigerinoides ruber и розовоокрашенная Globigerina rubescens (см. рис. 1). Во втором сверху Gs. ruber и наконец в верхнем горизонте присутствуют Gs. ruber, G. falconensis, Globorotalia scitula и ювенильная G. crassaformis (рис. 2). Самой северной точкой, в которой до сих пор отмечались представители розовоокрашенных Gs. ruber и G. rubescens , является скважина 410 DSDP (45°31’ с.ш.) в Атлантическом океане ( Былинская и др., 2002 ). Поэтому присутствие G. rubescens pink в колонке 2417 свидетельствует о значительно более тепловодных условиях по сравнению с современными или, скорее, о мощном усилении Северо-Атлантического течения, а именно его Западно-Шпицбергенской ветви в определенные моменты дегляциации. Альтернативное объяснение заключается в переотложении четвертичных тепловодных микроорганизмов, хотя его возможный источник в таких высоких широтах неизвестен.

Рис. 2. Тепловодные виды планктонных фораминифер из осадков колонки 2417.
Верхний ИВП в колонке 2417, характеризующийся обилием планктонных и умеренным количеством бентосных фораминифер, мы относим к нижней половине голоцена, а подстилающий его горизонт (35–110 см), практически лишенный микрофауны, к максимуму последнего оледенения (LGM). В силу этого, хотя есть много данных о том, что скорости морского осадконакопления в арктических районах во время оледенений сильно уменьшались ( Polyak et al., и др. ) (что видно и в нижней половине колонки), можно предположить среднюю скорость осадконакопления для рассматриваемого разреза примерно 3 см/1 тыс. лет. В этом случае нижний горизонт с тепловодной микрофауной соответствует последнему межледниковью, т. е. изотопно-кислородной стадии 5e. Этот вывод коррелируется с данными по колонке PS1535, взятой в проливе Фрама на такой же широте, на 4° западнее ( Spielhagen et al., 2004 ).
Почти все образцы колонки содержат зерна песчаной и гравийной размерности – материал ледового разноса (ice-rafted debris, IRD). Максимальный диаметр самых крупных зерен достигает 7–8 мм. Поскольку изученная колонка взята на глубине свыше 1500 м, то весь обломочный материал размерностью >250 µm очевидно поступал за счет приноса айсбергами и плавучими льдами. На рис. 1 показана кривая содержания IRD в колонке. И хотя она является предварительной, видно, что наибольшее содержание ледового обломочного материала в основном предшествует пикам обилия микропланктона или соответствует их нижней части, т. е. соответствует периодам дегляциации.
Бентосные фораминиферы в колонке 2417 представлены единым комплексом почти исключительно секреционных видов. Основным и наиболее многочисленным компонентом комплекса являются Cassidulina teretis и Oridorsalis tener . В образцах, соответствующих максимумам похолоданий, содержатся только единичные экземпляры C. teretis и иногда Rhabdammina sp. Примерно со средней части разреза тепловодные интервалы (197–228 см, 164–168 см, 16–33 см) характеризуются присутствием Cassidulina carinata , которая в отдельных образцах относительно многочисленна и даже замещает в комплексе C. teretis . Остальные бентосные фораминиферы представлены видами Triloculina tricarinata, Cassidulina reniforme, Islandiella norcrossi, Pullenia bulloides (в одном интервале), Cibicidoides wuellerstorfi (эпизодически), Epistominella exigua (в одном образце), Bulimina marginata, B. aculeata, Quinqueloculina aff . grandis, Pyrgo murrhina, Lagena sp ., Oolina sp ., Fissurina sp. Из агглютинирующих фораминифер редко встречаются Rhabdammina sp ., Reophax excentricus, Recurvoides sp ., Sigmoilopsis aff . schlumbergeri . В интервалах, соответствующих периодам дегляциации, содержатся шельфовые эльфидииды, принесенные плавучим льдом.
Читать дальшеИнтервал:
Закладка:
Похожие книги на «Строение и история развития литосферы»
Представляем Вашему вниманию похожие книги на «Строение и история развития литосферы» списком для выбора. Мы отобрали схожую по названию и смыслу литературу в надежде предоставить читателям больше вариантов отыскать новые, интересные, ещё непрочитанные произведения.












Обсуждение, отзывы о книге «Строение и история развития литосферы» и просто собственные мнения читателей. Оставьте ваши комментарии, напишите, что Вы думаете о произведении, его смысле или главных героях. Укажите что конкретно понравилось, а что нет, и почему Вы так считаете.