Николай Печуркин - Энергия и жизнь
Здесь есть возможность читать онлайн «Николай Печуркин - Энергия и жизнь» весь текст электронной книги совершенно бесплатно (целиком полную версию без сокращений). В некоторых случаях можно слушать аудио, скачать через торрент в формате fb2 и присутствует краткое содержание. Город: Новосибирск, Год выпуска: 1988, Издательство: Наука, сибирское отделение, Жанр: sci_biophys, на русском языке. Описание произведения, (предисловие) а так же отзывы посетителей доступны на портале библиотеки ЛибКат.

- Название:Энергия и жизнь
- Автор:
- Издательство:Наука, сибирское отделение
- Жанр:
- Год:1988
- Город:Новосибирск
- ISBN:нет данных
- Рейтинг книги:4 / 5. Голосов: 1
-
Избранное:Добавить в избранное
- Отзывы:
-
Ваша оценка:
Энергия и жизнь: краткое содержание, описание и аннотация
Предлагаем к чтению аннотацию, описание, краткое содержание или предисловие (зависит от того, что написал сам автор книги «Энергия и жизнь»). Если вы не нашли необходимую информацию о книге — напишите в комментариях, мы постараемся отыскать её.
Книга будет интересна для биологов, физиков, химиков, биофизиков, а также всех интересующихся общими вопросами развития.
Энергия и жизнь — читать онлайн бесплатно полную книгу (весь текст) целиком
Ниже представлен текст книги, разбитый по страницам. Система сохранения места последней прочитанной страницы, позволяет с удобством читать онлайн бесплатно книгу «Энергия и жизнь», без необходимости каждый раз заново искать на чём Вы остановились. Поставьте закладку, и сможете в любой момент перейти на страницу, на которой закончили чтение.
Интервал:
Закладка:
Н исп= Н пад— Н неисп. (1)
Сам поток энергии, захваченной системой, складывается из двух частей: доли, связанной с запасением энергии в биомассе — В , и потерь на организацию, поддержание и активность, т. е. расход энергии системой, — R :
Н исп= µ В — R [2] Опечатка в книге? Наверное: Н исп = µ В + R
. (2)
где µ — показатель прироста биомассы (или обновления в стационарном состоянии открытой системы).
Объединив (1) и (2), получим общее соотношение для потоков энергии в систему и ее трансформации в этой системе:
Н пад— Н неисп= Н исп= µ В + R (3)
Дадим формулировку энергетического принципа: в процессах развития надорганизменных систем (эволюции, экологических сукцессиях и перестройках) использованный биологической системой поток энергии Н исп возрастает, достигая локальных максимальных значений в стационарных состояниях.
Подчеркнем еще раз, что в формулировке данного принципа речь идет о стационарных состояниях, которые достигаются в процессе развития открытых биологических систем популяционного и более высоких уровней структурной организации, а также их эволюции.
Представим схематически наиболее предпочтительный тип изменения потоков энергии во времени, использованных системой (рис. 10,а). Здесь показаны и рост Н пад, т. е. рост захваченной энергии, и уменьшение Н неисп, т. е. снижение потерь. Штриховкой обозначено, что при этом происходит увеличение рассеяния энергии внутри системы Д с уменьшением трат на образование биомассы (это относится уже ко второму энергетическому принципу и будет обсуждаться позже). Естественно, что монотонное линейное увеличение Н падили снижение Н неиспне может иметь места в реальных системах. Гораздо типичнее скачкообразные переходы, связанные с качественными изменения в самой системе (рис. 10, б—г). Условия роста (или по крайней мере не убывания) потока использованной энергии Н испостаются для каждого момента времени.
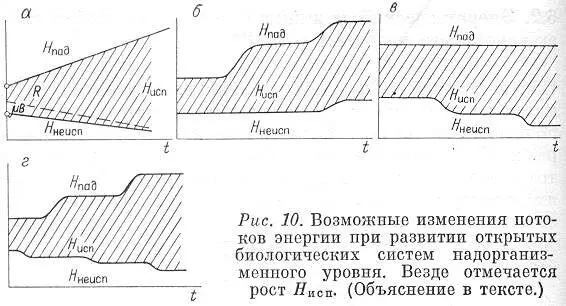
Рис. 10. Возможные изменения потоков энергии при развитии открытых биологических систем надорганизменного уровня. Везде отмечается рост Н исп. (Объяснение в тексте)
6.3. Экспериментальные эволюционные машины (ЭЭМ)
В предыдущем параграфе мы упоминали о том, что непрерывный рост микробных популяций в проточных системах позволяет экспериментально изучать микроэволюционные переходы, т.е. шаги эволюции.
Суть непрерывного процесса заключается в постоянной подаче питания в зону развития популяции и одновременном отборе избыточной части из рабочего пространства. Природная популяция, таким образом, осуществляя непрерывный обмен веществом с внешней средой и получая энергию извне, развивается в открытой системе, что является главной чертой ее динамики. В лабораторных условиях такой обмен осуществляется с помощью непрерывного потока питающей среды в рабочий объем ферментера и соответствующего оттока культуральной жидкости.
В классических периодических процессах нелимитированный рост популяции отмечается лишь во время экспоненциальной фазы, которая сменяется фазой замедления, т. е. торможения роста. При непрерывном культивировании можно застабилизировать рост популяции в любой точке на восходящей ветви S-образного роста популяции, в том числе и в экспоненциальной фазе. Для этого, как уже говорилось, необходимо непрерывно подавать свежую питательную среду для популяции и удалять избыточную часть популяции из рабочего объема. Для поддержания плотности популяции в заданной точке фазы нелимитированного роста применяются различные способы управления скоростью протока. Основное их свойство — наличие обратной связи между приростом концентрации биомассы и удалением части популяции из ферментера. Эти величины должны быть равными и это равенство, а с ним и концентрация биомассы — поддерживаются с помощью автоматических измеряющих и следящих устройств. Среди таких способов первым был турбидостат, в котором концентрация клеток поддерживается на определенном уровне за счет регулирования оптической плотности культуры. Применение его ограничено работой с оптически однородными средами.
Для процессов культивирования, в которых имеется прямая связь между приростом биомассы и изменением рН-культуры (например, при потреблении физиологически кислого источника азота), разработан и используется рН-статный способ управления скоростью протока. При этом скорость протока с помощью автоматических устройств уравнивается со скоростью изменения рН растущей популяцией, а следовательно, и со скоростью роста, что обеспечивает поддержание концентрации биомассы на заданном уровне.
Читать дальшеИнтервал:
Закладка:
Похожие книги на «Энергия и жизнь»
Представляем Вашему вниманию похожие книги на «Энергия и жизнь» списком для выбора. Мы отобрали схожую по названию и смыслу литературу в надежде предоставить читателям больше вариантов отыскать новые, интересные, ещё непрочитанные произведения.












Обсуждение, отзывы о книге «Энергия и жизнь» и просто собственные мнения читателей. Оставьте ваши комментарии, напишите, что Вы думаете о произведении, его смысле или главных героях. Укажите что конкретно понравилось, а что нет, и почему Вы так считаете.