А. Крылов - Регрессия как этап развития
Здесь есть возможность читать онлайн «А. Крылов - Регрессия как этап развития» — ознакомительный отрывок электронной книги совершенно бесплатно, а после прочтения отрывка купить полную версию. В некоторых случаях можно слушать аудио, скачать через торрент в формате fb2 и присутствует краткое содержание. Город: Москва, Год выпуска: 2017, ISBN: 2017, Издательство: Литагент Когито-Центр, Жанр: psy_generic, psy_theraphy, на русском языке. Описание произведения, (предисловие) а так же отзывы посетителей доступны на портале библиотеки ЛибКат.

- Название:Регрессия как этап развития
- Автор:
- Издательство:Литагент Когито-Центр
- Жанр:
- Год:2017
- Город:Москва
- ISBN:978-5-9270-0354-9
- Рейтинг книги:4 / 5. Голосов: 1
-
Избранное:Добавить в избранное
- Отзывы:
-
Ваша оценка:
Регрессия как этап развития: краткое содержание, описание и аннотация
Предлагаем к чтению аннотацию, описание, краткое содержание или предисловие (зависит от того, что написал сам автор книги «Регрессия как этап развития»). Если вы не нашли необходимую информацию о книге — напишите в комментариях, мы постараемся отыскать её.
Регрессия как этап развития — читать онлайн ознакомительный отрывок
Ниже представлен текст книги, разбитый по страницам. Система сохранения места последней прочитанной страницы, позволяет с удобством читать онлайн бесплатно книгу «Регрессия как этап развития», без необходимости каждый раз заново искать на чём Вы остановились. Поставьте закладку, и сможете в любой момент перейти на страницу, на которой закончили чтение.
Интервал:
Закладка:
Таким образом, было установлено, что под воздействием алкоголя меняется число нейрогенетически активных нейронов, вовлеченных в процесс приобретения нового опыта. В частности, меньше нейронов вовлекалось в этот процесс в тех областях, нейроны которых были вовлечены в приобретение первого навыка. Можно предположить, что под воздействием алкоголя дифференциация предыдущего опыта, связанная с аккомодационной реконсолидацией, происходит в меньшей степени, чем без такого воздействия. Возможно, вовлечение нейронов в новое обучение зависит от предыдущей истории их электрической активности (Guzowski et al., 2006) и связано с повышенной возбудимостью мембраны (Silva et al., 2009): поскольку позднее формируемый опыт реактивируется с большей вероятностью, чем более ранний (Сварник, 2016), это может приводить к снижению вероятности вовлечения элементов предыдущего опыта при формировании последующего.
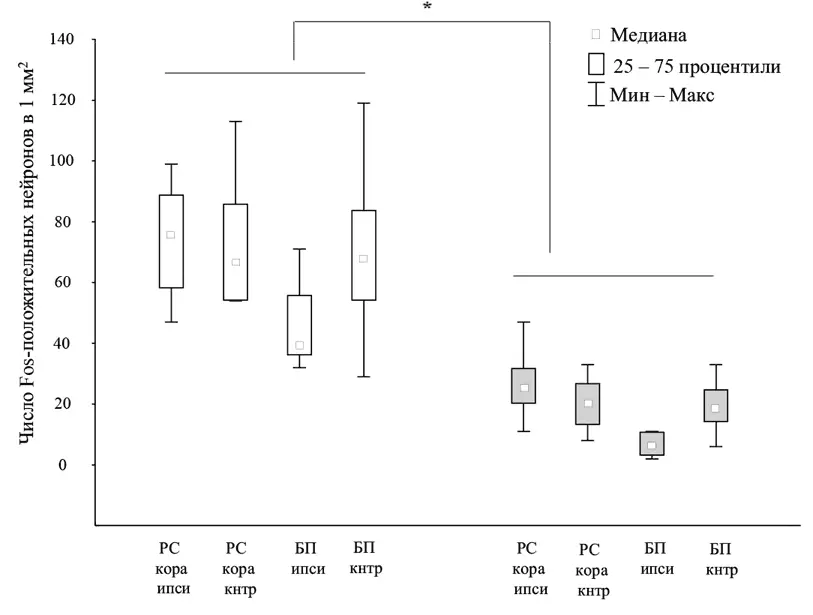
Рис. 3.Распределение нейрогенетических изменений (оцениваемых по белку Fos) в ретросплениальной коре (РС кора, RSA) и бочонковом поле соматосенсорной коры (БП, S1BF) головного мозга крыс при формировании нового навыка без введения алкоголя (белые столбики) и на фоне введения алкоголя (серые столбики), ипси – то же полушарие головного мозга по отношению к использованной в предыдущем опыте вибриссной подушке, кнтр – противоположное полушарие
Известно, что алкоголь может оказывать различное фармакологическое действие. Предполагается, что алкоголь действует на мембранные белки нейрона, в том числе и белки ионных каналов (Peoples et al., 1996), что приводит к подавлению его активности. Недавние исследования показывают, что эффекты алкоголя на нейронную активность варьируют в зависимости от того, в какой структуре находятся нейроны (White et al., 2000). Таким образом, нельзя говорить об общем глобальном влиянии алкоголя на генерацию потенциалов действия: в одних структурах показано увеличение активности нейронов (Huang et al., 2012), а в других, наоборот, подавление активности при введении алкоголя (Verbanck et al., 1990). В наших работах было показано, что введение алкоголя в большей степени подавляет активность нейронов, специализированных относительно вновь приобретенного опыта, причем известно, что доля таких нейронов выше в корковых областях (Alexandrov et al., 1990). В то же время амнестический эффект алкоголя главным образом связывается с подавлением активности NMDA-рецепторов и потенциированием ГАМК-опосредованного ингибирования активности нейронов (Nomura, Matsuki, 2008). Характерно, что плотность распределения NMDA-рецепторов по структурам мозга не одинакова (Petralia et al., 1994), в частности, плотность данных рецепторов выше в верхних слоях коры по сравнению с нижними (Conti et al., 1997), что может быть одним из факторов, лежащих в основе неоднородного влияния алкоголя на генерацию потенциалов действия и более выраженного подавления алкоголем активности специализированных нейронов в верхних слоях коры по сравнению с нижними (Alexandrov et al., 1990).
Экспрессия c-Fos не отражает напрямую повышенную активность нейронов, а маркирует скорее процесс рассогласования, возникающий в нейронах при нехватке у них различных метаболитов, необходимых им для выживания. Формирование функциональной системы, по-видимому, – это возможность для нейронов специализироваться в отношении данной системы, обеспечивать достижение результата на уровне целостного организма и тем самым устранять рассогласование, получая необходимые метаболиты от других клеток организма. Показано, что само по себе введение алкоголя незначительно изменяет число Fos-положительных клеток (Ryabinin et al., 1995; Segovia et al., 2013), однако в стрессовой ситуации число Fos-положительных нейронов снижается по сравнению с «безалкогольным» контролем. Таким образом, снижение числа Fos-положительных нейронов при обучении под воздействием алкоголя может означать снижение числа нейронов, находящихся в состоянии рассогласования и готовых вовлекаться в процессы системогенеза. Образно говоря, алкоголь, подавляя активность NMDA-рецепторов у нейронов, заставляет нейроны временно «забыть» о нужных им метаболитах. Интересно, что нарушение функции NMDA-рецепторов связывают с нарушением процессов рассогласования, оцениваемых, например, по величине негативности рассогласования (Chitty et al., 2015).
Полученные результаты позволяют сделать вывод о том, что при научении в условиях алкогольной регрессии в меньшей степени выражено вовлечение корковых областей в формирование новых элементов опыта . Это вовлечение основано на селекции активированных нейронов для последующей их специализации (см.: Александров, Сварник, 2009) в отношении формирующегося элемента. А селекция, как отмечал еще У. Джеймс, является основой формирования памяти (James, 1890). В условиях алкогольной регрессии также менее выражена реорганизация, «подстройка» уже имевшегося у индивида опыта, которая обеспечивает взаимосогласование раннее образованных и вновь сформированных систем.
Читать дальшеИнтервал:
Закладка:
Похожие книги на «Регрессия как этап развития»
Представляем Вашему вниманию похожие книги на «Регрессия как этап развития» списком для выбора. Мы отобрали схожую по названию и смыслу литературу в надежде предоставить читателям больше вариантов отыскать новые, интересные, ещё непрочитанные произведения.











Обсуждение, отзывы о книге «Регрессия как этап развития» и просто собственные мнения читателей. Оставьте ваши комментарии, напишите, что Вы думаете о произведении, его смысле или главных героях. Укажите что конкретно понравилось, а что нет, и почему Вы так считаете.