Вячеслав Мордкович - Основы биогеографии
Здесь есть возможность читать онлайн «Вячеслав Мордкович - Основы биогеографии» — ознакомительный отрывок электронной книги совершенно бесплатно, а после прочтения отрывка купить полную версию. В некоторых случаях можно слушать аудио, скачать через торрент в формате fb2 и присутствует краткое содержание. Город: Москва, Год выпуска: 2005, ISBN: 2005, Жанр: Прочая научная литература, geography_book, Детская образовательная литература, на русском языке. Описание произведения, (предисловие) а так же отзывы посетителей доступны на портале библиотеки ЛибКат.

- Название:Основы биогеографии
- Автор:
- Жанр:
- Год:2005
- Город:Москва
- ISBN:5-87317-189-0
- Рейтинг книги:5 / 5. Голосов: 1
-
Избранное:Добавить в избранное
- Отзывы:
-
Ваша оценка:
Основы биогеографии: краткое содержание, описание и аннотация
Предлагаем к чтению аннотацию, описание, краткое содержание или предисловие (зависит от того, что написал сам автор книги «Основы биогеографии»). Если вы не нашли необходимую информацию о книге — напишите в комментариях, мы постараемся отыскать её.
Основы биогеографии — читать онлайн ознакомительный отрывок
Ниже представлен текст книги, разбитый по страницам. Система сохранения места последней прочитанной страницы, позволяет с удобством читать онлайн бесплатно книгу «Основы биогеографии», без необходимости каждый раз заново искать на чём Вы остановились. Поставьте закладку, и сможете в любой момент перейти на страницу, на которой закончили чтение.
Интервал:
Закладка:
С позиций уровня взаимодействия живых существ между собой и с абиотической средой в структуре биогеографии выделяют географию биологических таксонов (ареалогию), географию флор, фаун, биот (флористику, фаунистику), географию экосистем и, наконец, географию ноосистем (от гр. “nous” – разум), т. е. систем жизнеобеспечения, созданных человеческим интеллектом заново или с использованием диких природных элементов. Ареалогия исследует пределы, форму, протяжённость территории распространения отдельных таксонов биоты. Задачей ареалогии является выделение, описание, объяснение и типологизация (выделение типов) ареалов, но без объяснения функциональных связей обладателей этих ареалов. Флористика и фаунистика занимаются выявлением и классификацией флор и фаун, т. е. совокупностей таксонов, имеющих ареалы со сходными параметрами. Характер уживаемости видов в рамках одной флоры или фауны остаётся за скобками биогеографического анализа. Наконец, географию экосистем, в отличие от экологии, интересуют не сами по себе функциональные взаимоотношения компонентов экосистемы, а закономерности размещения отдельных функциональных параметров или целых сообществ, биоценозов и биогеоценозов в пространстве.
География ноосистем специализируется на изучении обязательных зависимостей творений человеческого разума от существующей инфраструктуры земного пространства.
1.4. Основные теоретические позиции современной биогеографии
В современной биогеографии сосуществуют, по крайней мере, четыре теоретических направления.
Первое – это дисперсионная биогеография. Она базируется на дарвиновской концепции монофилетического происхождения таксонов филогенетической системы (каждый – из одного корня). Любой таксон имеет своё характерное время и место происхождения. То и другое устанавливается путем разделения и расстановки по древности рецентных, т. е. ныне живущих, и ископаемых представителей того или иного таксона. На этой основе реконструируются траектории его расселения по планете.
Особенности географического распространения таксона с позиций обсуждаемой теории предопределены адаптациями к перемещению, характерными для его представителей. Поскольку транспортные возможности у организмов из разных таксонов сильно различаются, то равномерного распределения жизни по земной поверхности не получается. В разных её частях складываются неодинаковые совокупности живых форм, которые можно лишь формально называть фаунами, флорами и т. д., ограничивая их содержание инвентарным списком видов, родов и т. д. Целью дисперсионной биогеографии служит описание, объяснение и классификация географических ареалов как можно большего числа таксонов, в перспективе желательно всех (рис. 7).
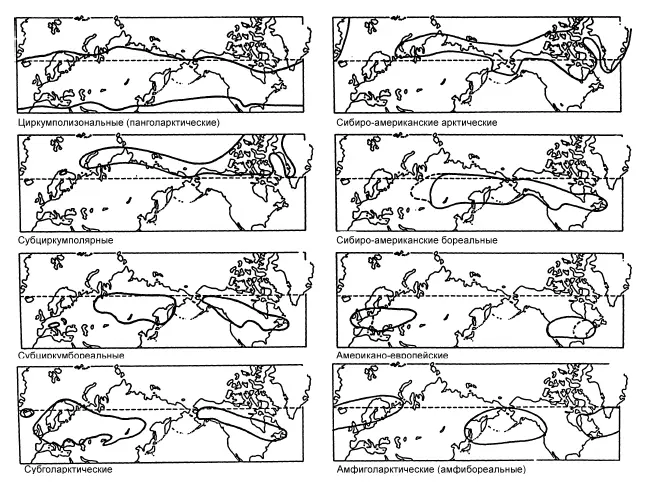
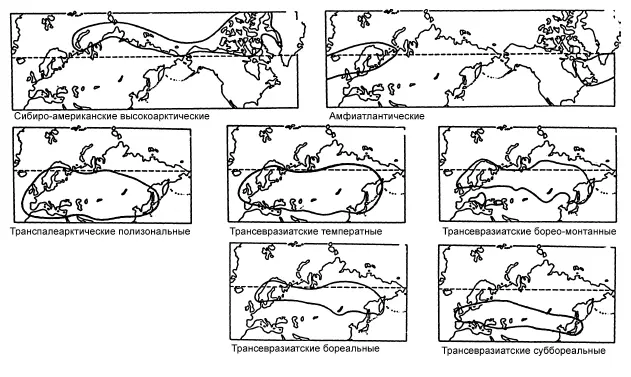
Рис. 7.Сравнение ареалов – основа дисперсионной биогеографии. По: Городков, 1986.
Императивом дисперсионной биогеографии является убеждение, что биогеографическую картину организмы формируют случайным образом, оперируя лишь собственными возможностями расселения, при пассивной роли среды.
Другим очень популярным теоретическим направлением биогеографии является флористико-фаунистическое. В противовес предыдущему, оно акцентирует внимание на формировании ограниченного набора очагов жизни, чётко ориентированных в географическом пространстве. В каждом из них исторически давно и непрерывно генерируются и интегрируются в совместимые и специфические совокупности таксоны животных и растений с ареалами более или менее одинаковой протяжённости, формы и инфраструктуры. Именно их следует именовать флорами и фаунами. Из этих очагов происходит дальнейшее расселение организмов. Выявление флор и фаун “де-факто” происходит не только путём статистического сравнения списков видов, но и выделения руководящих таксонов, типичных для разных очагов. Не менее важным критерием служит предварительное ограничение территорий и их разделение по специфике условий жизни. Эта специфика предопределена наличием физических преград (барьеров), препятствующих как вселению, так и выселению биоты с определённых территорий. Барьеры, мешая свободному распространению организмов даже с хорошими расселительными возможностями, создают режим изоляции внутри ограниченного пространства и способствуют там естественному отбору специфических форм жизни. Благодаря барьерам, реальные ареалы многих таксонов значительно меньше потенциальных. Из-за неоднородности экологического фона в пространстве у многих таксонов случаются разрывы ареалов. У истоков этих гипотез, объясняющих данные постулаты, стоял А. Уоллес (гипотеза оттеснённых реликтов и другие). Таким образом, суммарная биогеографическая картина мира по флористико-фаунистической теории складывается под влиянием не случайного, как в дисперсионной биогеографии, а детерминированного распределения и комбинирования таксонов, адекватно инфраструктуре пространства, заданной априори. Стандартная матрица пространства “диктует” одним и тем же таксонам совершенно разный порядок распределения в разных местах географической арены или одинаковое распределение разных таксонов в определённом месте часто вопреки их биологическому потенциалу к расселению. В противовес дисперсионной биогеографии – флористико-фаунистическую биогеографию можно ещё назвать интеграционной. А её императивом становится признание того, что биогеографическая картина мира является следствием не только биологического потенциала организмов к расселению, но и детерминированности географической арены. Её структура диктует порядок интеграции жизни в устойчивые совокупности таксонов флоры, фауны и т. и., характерные по составу (рис. 8).
Читать дальшеИнтервал:
Закладка:
Похожие книги на «Основы биогеографии»
Представляем Вашему вниманию похожие книги на «Основы биогеографии» списком для выбора. Мы отобрали схожую по названию и смыслу литературу в надежде предоставить читателям больше вариантов отыскать новые, интересные, ещё непрочитанные произведения.












Обсуждение, отзывы о книге «Основы биогеографии» и просто собственные мнения читателей. Оставьте ваши комментарии, напишите, что Вы думаете о произведении, его смысле или главных героях. Укажите что конкретно понравилось, а что нет, и почему Вы так считаете.