Александр Уголев - Естественные технологии биологических систем
Здесь есть возможность читать онлайн «Александр Уголев - Естественные технологии биологических систем» весь текст электронной книги совершенно бесплатно (целиком полную версию без сокращений). В некоторых случаях можно слушать аудио, скачать через торрент в формате fb2 и присутствует краткое содержание. Город: Ленинград, Год выпуска: 1987, Издательство: Наука, Жанр: Прочая научная литература, Медицина, Биология, на русском языке. Описание произведения, (предисловие) а так же отзывы посетителей доступны на портале библиотеки ЛибКат.

- Название:Естественные технологии биологических систем
- Автор:
- Издательство:Наука
- Жанр:
- Год:1987
- Город:Ленинград
- ISBN:нет данных
- Рейтинг книги:5 / 5. Голосов: 1
-
Избранное:Добавить в избранное
- Отзывы:
-
Ваша оценка:
Естественные технологии биологических систем: краткое содержание, описание и аннотация
Предлагаем к чтению аннотацию, описание, краткое содержание или предисловие (зависит от того, что написал сам автор книги «Естественные технологии биологических систем»). Если вы не нашли необходимую информацию о книге — напишите в комментариях, мы постараемся отыскать её.
Естественные технологии биологических систем — читать онлайн бесплатно полную книгу (весь текст) целиком
Ниже представлен текст книги, разбитый по страницам. Система сохранения места последней прочитанной страницы, позволяет с удобством читать онлайн бесплатно книгу «Естественные технологии биологических систем», без необходимости каждый раз заново искать на чём Вы остановились. Поставьте закладку, и сможете в любой момент перейти на страницу, на которой закончили чтение.
Интервал:
Закладка:
Вопрос.Может ли открытие неизвестных в настоящее время функциональных блоков существенно изменить представления о технологии и структурнофункциональной организации биологических систем?
Ответ.Как было отмечено ранее, некоторые недавно открытые функциональные блоки представляют собой комбинации блоков уже известных типов. Но нельзя исключить, что открытие неизвестных до сих пор функциональных блоков повлияет на понимание процессов, реализуемых живыми системами. Ярким примером служит белок мембраны эритроцитов, получивший на основании электрофоретических характеристик название «полоса 3» (П-3). Недавно сделан детальный обзор структуры и функций этого белка (Biochim. biophys. acta. 1986. Vol. 864. P. 145—167). Такой функциональный блок привлек внимание прежде всего в связи с тем, что он составляет около 25 % общего количества белка мембраны эритроцитов и присутствует в одной клетке в виде миллионов копий. Он обладает молекулярной массой 90 000—100 000 дальтон, пронизывает мембрану и имеет сложную внутриклеточную часть, или домен (рис. 47 и 48). Недавно стали понятны его основные функции, точнее, группы функций. Так, трансмембранная часть этого блока катализирует обмен неорганических анионов (особенно Сl -и НСО 3 -) через мембрану, а также участвует в транспорте воды. Далее, этот блок содержит антигенные детерминанты, в том числе в группах кровп А, В и О, а также резус-антиген. По всей вероятности, ой важен для опознавания стареющих и ненормальных клеток. Наконец, еще одна исключительно важная функция — связывание различных цитоплазматических белков, в том числе белков клеточного скелета.
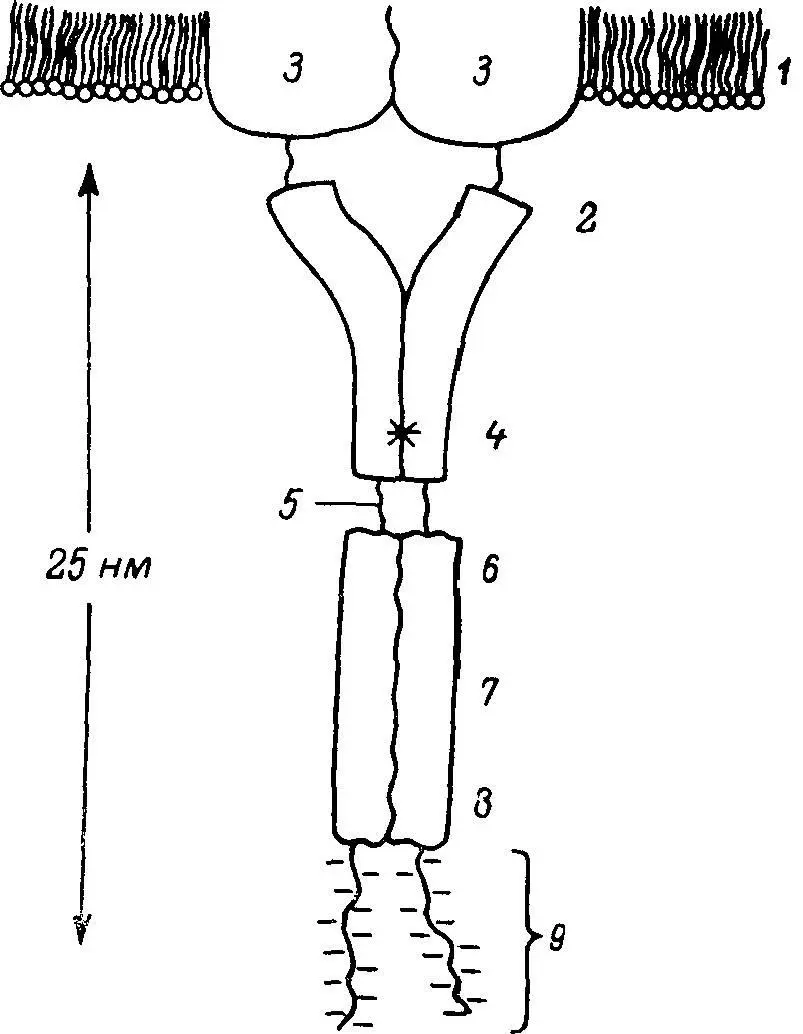
Рис. 48. Схема цитоплазматической части, или домена, белка «полоса 3» мембраны эритроцита человека.
1 — мембрана; 2 — цитоплазма; 3 — белок «полоса 3»; 4 — цистеиновый кластер; 5 — регулируемая область; 6 — участок связывания анкирина; 7 — участок (участки) связывания иммуноглобулина G; 8 — триптофановый кластер; 9 — ферменты, тельца Хейнца, участок связывания гемоглобина.
Мы обратили внимание лишь на некоторые стороны этой принципиально важной группы функций, которую можно было бы обозначить как цепторную функцию, а функциональный блок, выполняющий ее, отнести к группе цепторов.
Внутриклеточный домен белка П-3 благодаря функции связывания обеспечивает пространственную организацию многих функций мембраны и клетки в целом, структурные связи внутри нее и т.д. В частности, П-3, по-видимому, связывает многие ферменты, участвующие в гликолитическом цикле, и тем самым обеспечивает: 1) пространственную организацию цикла, 2) взаимодействие с субстратами, поступающими через мембрану, 3) эффективную передачу АТФ на АТФ-энергизируемые насосы. Все это крайне важно, так как ранее считалось, что большинство ферментов гликолитической системы пространственно не организовано. Точно так же до сих пор не было ясно, почему именно энергия гликолиза особенно необходима для реализации активного транспорта. Теперь можно полагать, что такая связь обусловлена пространственной и функциональной интеграцией гликолитических ферментов с мембранными АТФазами. Поэтому уже не кажется удивительным, что в клетках почечных канальцев П-3 локализован в базолатеральной мембране.
Однако «функциональные» соображения и данные относительно локализации некоторых насосов, использующих энергию АТФ, в апикальной мембране позволяют предположить, что в последней также будет обнаружен П-3. Особенно большое количество этого белка скорее всего может быть выявлено в обкладоч-ных клетках желудка, в апикальной мембране которых находятся мощные протонные насосы, обеспечивающие выделение в полость этого органа соляной кислоты. Ясно также, что П-3, первоначально интерпретированный как специфический белок мембраны эритроцитов, в действительности представляет собой универсальный блок, как эго было показано в прямых экспериментах.
Следующая важная функция внутриклеточного домена П-3 — связывание с белком 4.1 (полоса 4.1 при электрофорезе) и анкирином. Оба эти белка служат посредниками во взаимодействии основных элементов цитоскелета — актина и спектрина с клеточной мембраной. Эта функция определяет многие функциональные и структурные свойства клетки, в частности ее форму.
Наконец, для понимания функций эритроцитов существенна способность П-3 заякоривать вблизи мембраны молекулы гемоглобина. При этом сродство окисленного и восстановленного гемоглобина к П-3 различно. В целом наличие якорной функции у П-3 значительно меняет представления о многих свойствах клеток и в особенности о свойствах внутренней поверхности клеточной мембраны.
Читать дальшеИнтервал:
Закладка:
Похожие книги на «Естественные технологии биологических систем»
Представляем Вашему вниманию похожие книги на «Естественные технологии биологических систем» списком для выбора. Мы отобрали схожую по названию и смыслу литературу в надежде предоставить читателям больше вариантов отыскать новые, интересные, ещё непрочитанные произведения.












Обсуждение, отзывы о книге «Естественные технологии биологических систем» и просто собственные мнения читателей. Оставьте ваши комментарии, напишите, что Вы думаете о произведении, его смысле или главных героях. Укажите что конкретно понравилось, а что нет, и почему Вы так считаете.