Виктор Сбойчаков - Микробиология с основами эпидемиологии и методами микробиологических исследований
Здесь есть возможность читать онлайн «Виктор Сбойчаков - Микробиология с основами эпидемиологии и методами микробиологических исследований» — ознакомительный отрывок электронной книги совершенно бесплатно, а после прочтения отрывка купить полную версию. В некоторых случаях можно слушать аудио, скачать через торрент в формате fb2 и присутствует краткое содержание. ISBN: , Жанр: Медицина, Медицина, Прочая научная литература, Медицина, на русском языке. Описание произведения, (предисловие) а так же отзывы посетителей доступны на портале библиотеки ЛибКат.

- Название:Микробиология с основами эпидемиологии и методами микробиологических исследований
- Автор:
- Жанр:
- Год:неизвестен
- ISBN:978-5-299-00404-5
- Рейтинг книги:4 / 5. Голосов: 1
-
Избранное:Добавить в избранное
- Отзывы:
-
Ваша оценка:
Микробиология с основами эпидемиологии и методами микробиологических исследований: краткое содержание, описание и аннотация
Предлагаем к чтению аннотацию, описание, краткое содержание или предисловие (зависит от того, что написал сам автор книги «Микробиология с основами эпидемиологии и методами микробиологических исследований»). Если вы не нашли необходимую информацию о книге — напишите в комментариях, мы постараемся отыскать её.
Книга также будет полезна практическим лаборантам и фельдшерам, врачам микробиологам и студентам медицинских вузов.
Микробиология с основами эпидемиологии и методами микробиологических исследований — читать онлайн ознакомительный отрывок
Ниже представлен текст книги, разбитый по страницам. Система сохранения места последней прочитанной страницы, позволяет с удобством читать онлайн бесплатно книгу «Микробиология с основами эпидемиологии и методами микробиологических исследований», без необходимости каждый раз заново искать на чём Вы остановились. Поставьте закладку, и сможете в любой момент перейти на страницу, на которой закончили чтение.
Интервал:
Закладка:
При молочнокислом брожении (стрептококки, лактобациллы) водород переносится на пировиноградную кислоту и образуется молочная кислота:
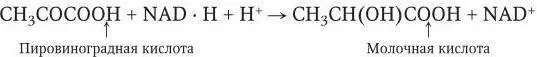
При спиртовом брожении (сахаромицеты) пировиноградная кислота сначала декарбоксилируется, т. е. от нее отщепляется СО 2,азатем промежуточный продукт (ацетальдегид) восстанавливается в этиловый спирт (этанол) в результате переноса водорода:
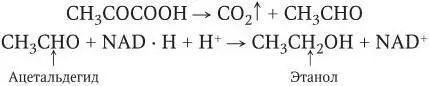
При маслянокислом брожении (клостридии) пировиноградная кислота превращается в ацетат [C H 3−C=0], связанный с коферментом (ацетил-СоА). Два таких остатка соединяются в ацетоацетат, который восстанавливается до н- масляной кислоты (СН 3СН 2СН 2СООН).
В 30-е годы XX века В. А. Энгельгардом, О. Варбургом, Ф. Лимпаном и Д. Диккенсаном было доказано, что кроме гликолиза в бактериальных клетках существует еще один путь расщепления углеводов – ступенчатый окислительный распад гексоз до пентоз и других сахаров с более короткой цепью. Ключевую роль в таких реакциях играют пентозофосфаты, в связи с чем этот цикл называется пентозофосфатным.
Суммарное уравнение этого цикла имеет вид:
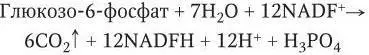
Небольшое число микроорганизмов, к которым относятся главным образом бактерии из рода Pseudomonas , получают энергию с помощью специфического метаболического пути – Энтнера – Дудорова. Существует предположение, что он появился в связи с высокой потребностью прокариот в пировиноградной кислоте как кратчайшему пути ее образования (всего 4 реакции, тогда как при гликолизе она образуется после 9 реакций).
В аэробных условиях пировиноградная кислота (ПВК) подвергается окислительному декарбоксилированию в цикле трикарбоновых кислот до конечных продуктов: СО 2иН 2О. Непосредственным реакциям цикла предшествует подготовительная фаза – окисление пировиноградной кислоты до ацетилкоэнзима А (ацетил = СоА):
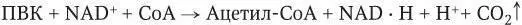
Окислительное декарбоксилирование ПВК катализируется пируватдегидрогеназной системой. Она состоит не менее чем из трех ферментов, которые используют пять коферментов: тиамидинфосфат, амид липоевой кислоты, коэнзим А, FAD, NAD.
При аэробном окислении 1 моля глюкозы клетка получает 38 молей АТФ, тогда как при гликолизе только два.
В цикле трикарбоновых кислот от субстратов отделяются протоны и электроны. Они поступают на коферменты NAD +и FAD +, которые передают их в дыхательную цепь, образованную окислительно-восстановительными ферментами, находящимися на плазматической мембране у прокариот и на внутренней мембране митохондрий у эукариот. Передвигаясь от одного переносчика электронов к другому, электроны опускаются на все более низкие энергетические уровни, отдавая порциями свою энергию. В последнем звене цепи они восстанавливают молекулярный кислород. Освобожденная при переносе электронов по дыхательной цепи энергия запасается в фосфатных связях АТФ. Окислительно-восстановительные ферменты представлены дегидрогеназами, флавопротеидами, цитохромами, а также убихиноном и белками, содержащими железо и серу.
Автотрофный метаболизм осуществляется бактериями путем фотосинтеза и хемосинтеза. Для облигатных автотрофов фотосинтез – единственный источник энергии: у них нет процессов диссимиляции, поставляющих АТФ. Фотосинтез состоит из двух фаз: световой и темновой. Световой процесс можно представить так:
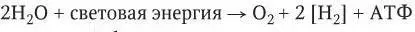
Во время темновой фазы атомы водорода, поставляемые световыми реакциями, используются для восстановления СО 2до углеводов согласно общему уравнению фотосинтеза:
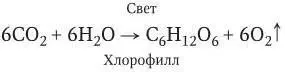
При этом на каждый моль синтезированного углевода запасается около 160 кДж энергии.
Русским микробиологом С. Н. Виноградским было показано, что хемосинтезирующие бактерии получают энергию при окислении неорганических соединений:
Читать дальшеИнтервал:
Закладка:
Похожие книги на «Микробиология с основами эпидемиологии и методами микробиологических исследований»
Представляем Вашему вниманию похожие книги на «Микробиология с основами эпидемиологии и методами микробиологических исследований» списком для выбора. Мы отобрали схожую по названию и смыслу литературу в надежде предоставить читателям больше вариантов отыскать новые, интересные, ещё непрочитанные произведения.












Обсуждение, отзывы о книге «Микробиология с основами эпидемиологии и методами микробиологических исследований» и просто собственные мнения читателей. Оставьте ваши комментарии, напишите, что Вы думаете о произведении, его смысле или главных героях. Укажите что конкретно понравилось, а что нет, и почему Вы так считаете.