Коллектив авторов - Общая вирусология с основами таксономии вирусов позвоночных
Здесь есть возможность читать онлайн «Коллектив авторов - Общая вирусология с основами таксономии вирусов позвоночных» — ознакомительный отрывок электронной книги совершенно бесплатно, а после прочтения отрывка купить полную версию. В некоторых случаях можно слушать аудио, скачать через торрент в формате fb2 и присутствует краткое содержание. Год выпуска: 2012, Издательство: Литагент БИБКОМ, Жанр: Медицина, Детская образовательная литература, на русском языке. Описание произведения, (предисловие) а так же отзывы посетителей доступны на портале библиотеки ЛибКат.

- Название:Общая вирусология с основами таксономии вирусов позвоночных
- Автор:
- Издательство:Литагент БИБКОМ
- Жанр:
- Год:2012
- ISBN:нет данных
- Рейтинг книги:5 / 5. Голосов: 1
-
Избранное:Добавить в избранное
- Отзывы:
-
Ваша оценка:
Общая вирусология с основами таксономии вирусов позвоночных: краткое содержание, описание и аннотация
Предлагаем к чтению аннотацию, описание, краткое содержание или предисловие (зависит от того, что написал сам автор книги «Общая вирусология с основами таксономии вирусов позвоночных»). Если вы не нашли необходимую информацию о книге — напишите в комментариях, мы постараемся отыскать её.
Общая вирусология с основами таксономии вирусов позвоночных — читать онлайн ознакомительный отрывок
Ниже представлен текст книги, разбитый по страницам. Система сохранения места последней прочитанной страницы, позволяет с удобством читать онлайн бесплатно книгу «Общая вирусология с основами таксономии вирусов позвоночных», без необходимости каждый раз заново искать на чём Вы остановились. Поставьте закладку, и сможете в любой момент перейти на страницу, на которой закончили чтение.
Интервал:
Закладка:
Покрытые вакуоли сливаются с другими, более крупными цитоплазматическими вакуолями, образуя рецептосомы, содержащие рецепторы, но не содержащие клатрин, а те в свою очередь сливаются с лизосомами. Таким путем проникшие в клетку белки обычно транспортируются в лизосомы, где происходит их распад на аминокислоты; они могут и миновать лизосомы, и накапливаться в других участках клетки в недеградированной форме. Альтернативой рецепторного эндоцитоза является жидкостный эндоцитоз, когда инвагинация происходит не в специализированных участках мембраны.
Большинство оболочечных и безоболочечных вирусов животных проникает в клетку по механизму рецепторного эндоцитоза. Эндоцитоз обеспечивает внутриклеточный транспорт вирусной частицы в составе эндоцитарной вакуоли, поскольку вакуоль может двигаться в любом направлении и сливаться с клеточными мембранами (включая ядерную мембрану), освобождая вирусную частицу в соответствующих внутриклеточных участках. Таким путем, например, ядерные вирусы попадают в ядро, а реовирусы – в лизосомы. Однако проникшие в клетку вирусные частицы находятся в составе вакуоли и отделены от цитоплазмы ее стенками. Им предстоит пройти ряд этапов, прежде чем они смогут вызвать инфекционный процесс.
Слияние вирусной и клеточной мембран. Для того чтобы внутренний компонент вируса мог пройти через клеточную мембрану, вирус использует механизм слияния мембран. У оболочечных вирусов слияние обусловлено точечным взаимодействием вирусного белка слияния с липидами клеточной мембраны, в результате которого вирусная липопротеидная оболочка интегрирует с клеточной мембраной, а внутренний компонент вируса оказывается по другую ее сторону. У безоболочечных вирусов один из поверхностных белков также взаимодействует с липидами клеточных мембран, в результате чего внутренний компонент проходит через мембрану. Большинство вирусов животных выходит в цитозол из рецептосомы.
Если при эндоцитозе вирусная частица является пассивным пассажиром, то при слиянии она становится активным участником процесса. Белком слияния является один из ее поверхностных белков. К настоящему времени этот белок идентифицирован лишь у парамиксовирусов и ортомиксовирусов. У парамиксовирусов этот белок (F-белок) представляет собой один из двух гликопротеидов, находящихся на поверхности вирусной частицы.
Функцию белка слияния у вируса гриппа выполняет малая гемагглютинирующая субъединица, НА2.
Парамиксовирусы вызывают слияние мембран при нейтральном рН, и внутренний компонент этих вирусов может проникать в клетку непосредственно через плазматическую мембрану. Однако большинство оболочечных и безоболочечных вирусов вызывают слияние мембран только при низком значении рН – от 5,0 до 5,75. Если к клеткам добавить слабые основания (хлорид аммония и др.), которые в эндоцитарных вакуолях повышают рН до 6,0, слияния мембран не происходит, вирусные частицы остаются в вакуолях, и инфекционный процесс не возникает. Строгая зависимость слияния мембран от значений рН обусловлена конформационными изменениями вирусных белков слияния.
В лизосоме постоянно имеется низкое значение рН (4,9). В эндоцитарной вакуоли (рецептосоме) закисление создается за счет АТФ-зависимого «протонового насоса» еще на клеточной поверхности при образовании покрытой вакуоли. Закисление эндоцитарной вакуоли имеет большое значение для проникающих в клетку физиологических лигандов, так как низкое значение рН способствует диссоциации лиганда от рецептора я рециркуляции рецепторов.
Тот же механизм, который лежит в основе слияния вирусных и клеточных мембран, обусловливает индуцированный вирусами гемолиз и слияние плазматических мембран, прилежащих друг к другу клеток с образованием многоядерных клеток, симпластов и синцитиев. Вирусы вызывают два типа слияния клеток: 1) «слияние снаружи» и 2) «слияние изнутри» (рисунок 20). «Слияние снаружи» происходит при высокой множественности инфекции и обнаруживается в течение первых часов после заражения.
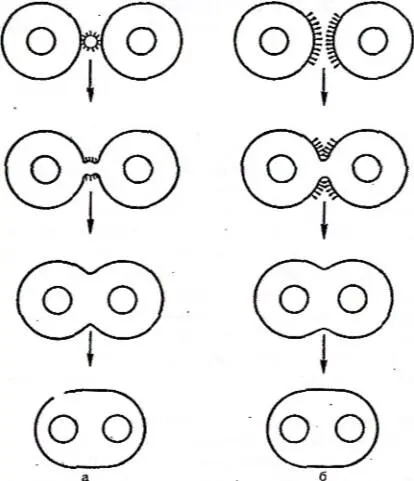
а – слияние извне; б – слияние изнутри.
Рисунок 20 – Слияние клеточных мембран при заражении клеток вирусом
Этот тип слияния, описанный для парамиксовирусов, обусловлен белками заражающего вируса и не требует внутриклеточного синтеза вирусных компонентов. Напротив, «слияние изнутри» происходит при низкой множественности инфекции, обнаруживается на сравнительно поздних стадиях инфекционного процесса и обусловлено вновь синтезированными вирусными белками. «Слияние изнутри» описано для многих вирусов: вирусов герпеса, онковирусов, возбудителей медленных инфекций и др. Этот тип слияния вызывают те же вирусные гликопротеиды, которые обеспечивают проникновение вируса в клетку.
Читать дальшеИнтервал:
Закладка:
Похожие книги на «Общая вирусология с основами таксономии вирусов позвоночных»
Представляем Вашему вниманию похожие книги на «Общая вирусология с основами таксономии вирусов позвоночных» списком для выбора. Мы отобрали схожую по названию и смыслу литературу в надежде предоставить читателям больше вариантов отыскать новые, интересные, ещё непрочитанные произведения.












Обсуждение, отзывы о книге «Общая вирусология с основами таксономии вирусов позвоночных» и просто собственные мнения читателей. Оставьте ваши комментарии, напишите, что Вы думаете о произведении, его смысле или главных героях. Укажите что конкретно понравилось, а что нет, и почему Вы так считаете.