Коллектив авторов - Общая вирусология с основами таксономии вирусов позвоночных
Здесь есть возможность читать онлайн «Коллектив авторов - Общая вирусология с основами таксономии вирусов позвоночных» — ознакомительный отрывок электронной книги совершенно бесплатно, а после прочтения отрывка купить полную версию. В некоторых случаях можно слушать аудио, скачать через торрент в формате fb2 и присутствует краткое содержание. Год выпуска: 2012, Издательство: Литагент БИБКОМ, Жанр: Медицина, Детская образовательная литература, на русском языке. Описание произведения, (предисловие) а так же отзывы посетителей доступны на портале библиотеки ЛибКат.

- Название:Общая вирусология с основами таксономии вирусов позвоночных
- Автор:
- Издательство:Литагент БИБКОМ
- Жанр:
- Год:2012
- ISBN:нет данных
- Рейтинг книги:5 / 5. Голосов: 1
-
Избранное:Добавить в избранное
- Отзывы:
-
Ваша оценка:
Общая вирусология с основами таксономии вирусов позвоночных: краткое содержание, описание и аннотация
Предлагаем к чтению аннотацию, описание, краткое содержание или предисловие (зависит от того, что написал сам автор книги «Общая вирусология с основами таксономии вирусов позвоночных»). Если вы не нашли необходимую информацию о книге — напишите в комментариях, мы постараемся отыскать её.
Общая вирусология с основами таксономии вирусов позвоночных — читать онлайн ознакомительный отрывок
Ниже представлен текст книги, разбитый по страницам. Система сохранения места последней прочитанной страницы, позволяет с удобством читать онлайн бесплатно книгу «Общая вирусология с основами таксономии вирусов позвоночных», без необходимости каждый раз заново искать на чём Вы остановились. Поставьте закладку, и сможете в любой момент перейти на страницу, на которой закончили чтение.
Интервал:
Закладка:
Вирусы обладают механизмами, которые позволяют прединициирующему комплексу эффективно избегать большей части 5’-НТР и начинать сканирование в пределах области инициирующего кодона AUG вирусной мРНК. Первая ассоциация Met-tRNAi с инициирующим кодоном AUG приводит к гидролизу GTP в тройном комплексе, связанные факторы инициирования освобождаются и 60S субъединица рибосомы присоединяется к прединициирующему комплексу. В результате 80S инициирующий комплекс опосредует стадию элонгации трансляции. В этой модели, инициация начинается с 5’-проксимального AUG кодона. Однако выбор кодона AUG для инициации трансляции зависит частично от его контекста, где каноническая accAUGg последовательность проявляет самую высокую активность к инициированию. Отступление от этой последовательности связано с так называемым негерметичным сканированием, в котором прединициирующий комплекс редко признает неканонический или слабый AUG и сканирует мимо, чтобы начать трансляцию с кодона, более соответствующего каноническому инициирующему AUG.
Негерметичное сканирование при инициации трансляции популярно среди вирусов, а у ретровирусов оно может обеспечивать определенные стехиометрические взаимоотношения продуктов трансляции.
Удлинение. В течение стадии элонгации трансляции мРНК связана со многими 80S рибосомами, или полисомами, поскольку аминокислотные остатки последовательно присоединяются к СООН-концу растущей цепи пептидов. Во многих вирусных системах жизненный цикл разграничен на ранние и поздние события, которые могут различаться дифференцированным привлечением вирусных мРНК в полисомные комплексы в определенное время после инфекции. Например, у вируса простого герпеса (HSV-1) это часто совпадает с синтезом факторов латентности и детерминант вирулентности. Процесс трансляции на стадии элонгации подчинен вирусной регуляции. Механизмы контроля элонгации включают, например, рибосомальный сдвиг рамки считывания и направленную вирусом модификацию EF-1. Первый механизм распространен у ретровирусов и связан с наличием дополнительных ORFs в пределах вирусной мРНК.
Терминация. Процесс завершения трансляции происходит в тот момент, когда 80S рибосома сталкивается в рамке считывания с терминирующим кодоном в пределах последовательности мРНК. Терминирующий кодон является фактором, который запускает процесс гидролиза связи пептидной цепи и тРНК, освобождает синтезированный полипептид от 80S рибосомы и разобщает субъединицы рибосомы. Как только завершение синтеза произошло, 40S субъединица может продолжить сканировать мРНК.
При считывании мультицистронной последовательности завершение трансляции может сопровождаться переинициированием трансляции ниже расположенного гена. Завершение трансляции переинициированием распространено среди вирусов и используется ими, чтобы управлять синтезом определенного генного продукта. После открытия полиаденилирования э-мРНК стало ясно, что поли-A трек играет важную роль в трансляции мРНК в клетках эукариот. Современные исследования показали, что определенную роль в стимулирующей функции поли-A трека на процесс трансляции играет поли-A-связывающий белок (PABP). В клетках животных PABP взаимодействует с элементами кэп-связывающего комплекса, включая в его состав 5’– конец мРНК и создавая, таким образом, трансляционный комплекс в форме «закрытой петли» (рисунок 17).
мРНК связующий EIF4F инициирующий комплекс взаимодействует с 3’– концом мРНК через PABP. Поли-A последовательность в пределах 3’-НТР прямым образом связывает PABP с мРНК. PABP добивается взаимодействия с кэпсвязывающим комплексом непосредственно через eIF4G (4G) или косвенно через взаимодействие eIF4G, eIF4A (4A) и Paip-1. Сборка комплекса замкнутой системы может стабилизировать взаимодействие 40S субъединицы рибосомы с мРНК.
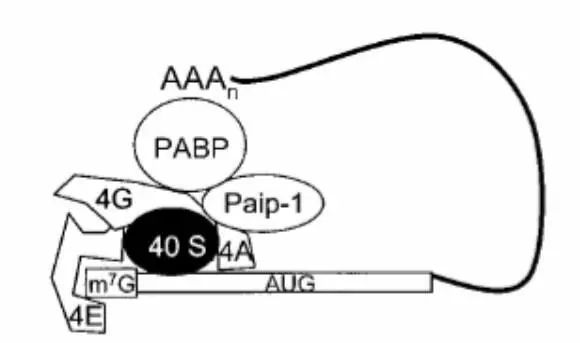
Рисунок 17 – Модель трансляционного комплекса в замкнутой системе с мРНК
PABP формирует закрытую петлю путем связывания eIF4G и белка Paip-1. Paip-1 взаимодействует с компонентами кэп-связывающего комплекса мРНК, включая eIF4G и eIF4A-хеликазу. Изучение инициации трансляции в дрожжах и растениях показали, что взаимодействие между PABP и eIF4G стимулирует трансляцию мРНК. Сближение концов мРНК, обеспеченное закрытым трансляционным комплексом, вносит вклад в стабильность мРНК и 5’-кэп-комплекса и обеспечивает эффективную сборку полирибосом. Таким образом, полный эффект закрытой петли заключается в увеличении эффективности трансляции. Вирусы используют закрытый трансляционный комплекс как средство переключения трансляционного аппарата клетки на трансляцию вирусных мРНК путем разрушения или модификации РАВР.
Читать дальшеИнтервал:
Закладка:
Похожие книги на «Общая вирусология с основами таксономии вирусов позвоночных»
Представляем Вашему вниманию похожие книги на «Общая вирусология с основами таксономии вирусов позвоночных» списком для выбора. Мы отобрали схожую по названию и смыслу литературу в надежде предоставить читателям больше вариантов отыскать новые, интересные, ещё непрочитанные произведения.












Обсуждение, отзывы о книге «Общая вирусология с основами таксономии вирусов позвоночных» и просто собственные мнения читателей. Оставьте ваши комментарии, напишите, что Вы думаете о произведении, его смысле или главных героях. Укажите что конкретно понравилось, а что нет, и почему Вы так считаете.