Коллектив авторов - Общая вирусология с основами таксономии вирусов позвоночных
Здесь есть возможность читать онлайн «Коллектив авторов - Общая вирусология с основами таксономии вирусов позвоночных» — ознакомительный отрывок электронной книги совершенно бесплатно, а после прочтения отрывка купить полную версию. В некоторых случаях можно слушать аудио, скачать через торрент в формате fb2 и присутствует краткое содержание. Год выпуска: 2012, Издательство: Литагент БИБКОМ, Жанр: Медицина, Детская образовательная литература, на русском языке. Описание произведения, (предисловие) а так же отзывы посетителей доступны на портале библиотеки ЛибКат.

- Название:Общая вирусология с основами таксономии вирусов позвоночных
- Автор:
- Издательство:Литагент БИБКОМ
- Жанр:
- Год:2012
- ISBN:нет данных
- Рейтинг книги:5 / 5. Голосов: 1
-
Избранное:Добавить в избранное
- Отзывы:
-
Ваша оценка:
Общая вирусология с основами таксономии вирусов позвоночных: краткое содержание, описание и аннотация
Предлагаем к чтению аннотацию, описание, краткое содержание или предисловие (зависит от того, что написал сам автор книги «Общая вирусология с основами таксономии вирусов позвоночных»). Если вы не нашли необходимую информацию о книге — напишите в комментариях, мы постараемся отыскать её.
Общая вирусология с основами таксономии вирусов позвоночных — читать онлайн ознакомительный отрывок
Ниже представлен текст книги, разбитый по страницам. Система сохранения места последней прочитанной страницы, позволяет с удобством читать онлайн бесплатно книгу «Общая вирусология с основами таксономии вирусов позвоночных», без необходимости каждый раз заново искать на чём Вы остановились. Поставьте закладку, и сможете в любой момент перейти на страницу, на которой закончили чтение.
Интервал:
Закладка:
В репликативных вилках одна нить (ведущая) копируется непрерывно в направлении от 5’– к 3’-концу. Поскольку другая нить (отстающая) должна также синтезироваться от 5’– к 3’-концу, она копируется с перерывами, многократно инициируя синтез и соединяя короткие фрагменты Оказаки. Синтез ДНК в репликативной вилке обеспечивается целым набором белков-ферментов, которые могут иметь разное происхождение. Мелкие ДНК-содержащие вирусы используют клеточные репликативные белки. Лучше всех изучена репликация полиомавируса SV40, где вовлеченные репликативные белки были идентифицированы в бесклеточной системе in vitro .
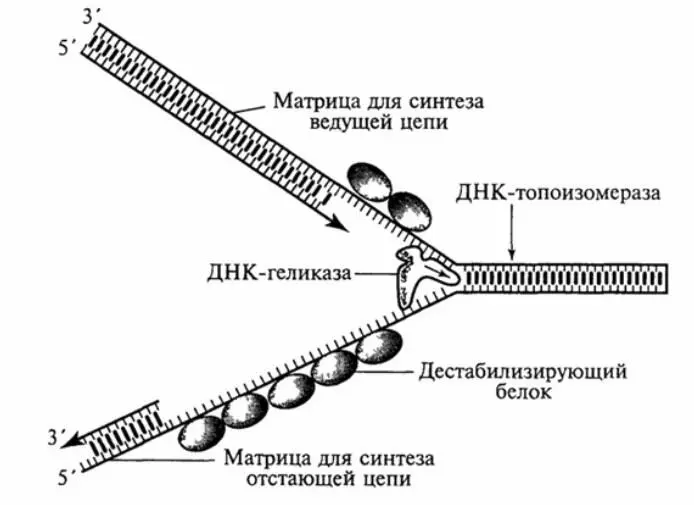
Рисунок 9 – Схема репликации ДНК с использованием репликативной вилки
Установлено, что в репликации ДНК SV40 принимают участие 10 белков. Девять из них имеют клеточное происхождение: ДНК-полимераза α (ответственна за инициацию синтеза ДНК в точке ori и синтез отстающей нити); праймаза (связана с ДНК-полимеразой и праймирует синтез фрагментов Оказаки); ДНК-полимераза d (ответственна за синтез лидирующей нити и завершение синтеза фрагментов Оказаки); пролиферативный клеточный ядерный антиген (PCNA), который связывается с ДНК-полимеразой d и формирует кольцо вокруг ДНК, увеличивая процессивность полимеразы; гетеропентамерный репликативный фактор C – RF-C (присоединяет кольцо PCNA на ДНК и стимулирует полимеразу d); RPA – онДНК-связывающий белок; РНаза H (удаляет все кроме одного рибонуклеотиды РНК-праймера); экзонуклеаза FEN-1, также известная как MF-1 (удаляет оставшейся рибонуклеотид); ДНК-лигаза I (лигирует фрагменты Оказаки); топоизомераза I и/или топоизомераза II (снимает сверхспирализацию в течение синтеза). Единственный вирусный белок, который требуется для репликации ДНК SV40 – это большой T-антиген, который обладает свойствами хеликазы и обеспечивает расплетение двунитевой структуры в репликативной вилке.
Другие вирусы сами обеспечивают почти все белки репликативной вилки. Например, фаза элонгации при репликации ДНК аденовируса в условиях in vitro обеспечивается одной аденовирусной субъединицей ДНК-полимеразы, аденовирусным однонитевым ДНК-связывающим белком, который может увеличивать процессивность полимеразы, и клеточной топоизомеразой I или II. Это простота частично связана с необычным характером репликации ДНК аденовируса, в которой отсутствует синтез отстающей цепи.
Крупные ДНК-вирусы еще в большей степени обеспечивают себя ферментами репликации. Например, вирусы герпеса кодируют ДНК-полимеразу, фактор элонгации, праймазо-хеликазный комплекс, однонитевой ДНК-связывающий белок и, вероятно, еще ряд вирусных белков, которые не идентифицированы.
Терминация синтеза. В случае кольцевых геномов окончание синтеза и расхождение геномов упрощены, поскольку синтез дочерней цепи идет по кругу и в конце полного оборота в точке ori или при двунаправленной репликации в середине кольца 3’– и 5’-концы вновь синтезированной цепи совмещаются и лигируются. Попарно сцепленные кольца разъединяются топоизомеразой. В линейных ДНК, синтезированных с помощью РНК-затравок, все обстоит сложнее. Удаление РНК-праймера дает молекулу ДНК с выступающим 3’-концом и пробелом на 5’-конце. Предложено два способа завершения репликации с образованием полной копии матричной цепи: с использованием конкатамеров или через образование шпильки.
Основные схемы репликации ДНК-геномных вирусов .
1 Терминальная инициация с помощью самозатравочного механизма.
2 Терминальная инициация с помощью белок-нуклеотидной (Б-Н) затравки.
3 Механизм катящегося кольца.
4 Схема Кернса.
5 Репликация через интеграцию.
1 Репликация с использованием терминальной инициации при помощи самозатравочного механизма (рисунок 10). Такой тип репликации геномной ДНК имеют парвовирусы, у которых геном представлен линейной онДНК, имеющей на обоих конца самокомплементарные последовательности, формирующие шпилечные структуры. 3’-конец ДНК имеет уникальную последовательность размером 125 нуклеотидов, образующую двунитевую Т-образную шпилечную структуру, которая играет роль затравки для ДНК-полимеразы.
ДНК-полимераза в результате репарационного синтеза комплементарной цепи воссоздает дуплекс, обе цепи которого на одном конце ковалентно соединены. При этом 3’-концевой сегмент родительского генома в качестве матрицы не используется. Следовательно, полного воспроизведения вирусного генома пока не произошло. На следующем этапе вирусоспецифический фермент вносит разрыв в родительскую цепь на границе между реплицированным и нереплицированным участками последовательности (между 125 и 126 нуклеотидами).
Читать дальшеИнтервал:
Закладка:
Похожие книги на «Общая вирусология с основами таксономии вирусов позвоночных»
Представляем Вашему вниманию похожие книги на «Общая вирусология с основами таксономии вирусов позвоночных» списком для выбора. Мы отобрали схожую по названию и смыслу литературу в надежде предоставить читателям больше вариантов отыскать новые, интересные, ещё непрочитанные произведения.












Обсуждение, отзывы о книге «Общая вирусология с основами таксономии вирусов позвоночных» и просто собственные мнения читателей. Оставьте ваши комментарии, напишите, что Вы думаете о произведении, его смысле или главных героях. Укажите что конкретно понравилось, а что нет, и почему Вы так считаете.