К.ПРИБРАМ - ЯЗЫКИ МОЗГА Экспериментальные парадоксы и принципы нейропсихологии
Здесь есть возможность читать онлайн «К.ПРИБРАМ - ЯЗЫКИ МОЗГА Экспериментальные парадоксы и принципы нейропсихологии» весь текст электронной книги совершенно бесплатно (целиком полную версию без сокращений). В некоторых случаях можно слушать аудио, скачать через торрент в формате fb2 и присутствует краткое содержание. Жанр: Медицина, на русском языке. Описание произведения, (предисловие) а так же отзывы посетителей доступны на портале библиотеки ЛибКат.

- Название:ЯЗЫКИ МОЗГА Экспериментальные парадоксы и принципы нейропсихологии
- Автор:
- Жанр:
- Год:неизвестен
- ISBN:нет данных
- Рейтинг книги:5 / 5. Голосов: 1
-
Избранное:Добавить в избранное
- Отзывы:
-
Ваша оценка:
ЯЗЫКИ МОЗГА Экспериментальные парадоксы и принципы нейропсихологии: краткое содержание, описание и аннотация
Предлагаем к чтению аннотацию, описание, краткое содержание или предисловие (зависит от того, что написал сам автор книги «ЯЗЫКИ МОЗГА Экспериментальные парадоксы и принципы нейропсихологии»). Если вы не нашли необходимую информацию о книге — напишите в комментариях, мы постараемся отыскать её.
Под редакцией и с предисловием
действительного члена АПН СССР
А. Р. Лурия
Издательство «Прогресс» Москва 1975
Редакция литературы по философии
Перевод на русский язык с изменениями. "Прогресс"
ЯЗЫКИ МОЗГА Экспериментальные парадоксы и принципы нейропсихологии — читать онлайн бесплатно полную книгу (весь текст) целиком
Ниже представлен текст книги, разбитый по страницам. Система сохранения места последней прочитанной страницы, позволяет с удобством читать онлайн бесплатно книгу «ЯЗЫКИ МОЗГА Экспериментальные парадоксы и принципы нейропсихологии», без необходимости каждый раз заново искать на чём Вы остановились. Поставьте закладку, и сможете в любой момент перейти на страницу, на которой закончили чтение.
Интервал:
Закладка:
Однако проблема речи еще не исчерпывается этим. Как мы показали в предыдущих двух главах, приматы могут передавать сообщения относительно окружающего их мира и относительно своего внутреннего мира с помощью примитивных знаков и символов. Но у приматов не существует языковой структуры. Поэтому возникает вопрос: какие же изменения должны были произойти, чтобы человеческий мозг стал таким, каков он есть, то-есть чтобы стало возможным существование языковых систем? Отсюда следует другой вопрос: почему владение языком представляет собой такое мощное орудие адаптации человека и прогресса человечества? В следующих разделах мы рассмотрим эти вопросы.
На вопрос о том, что делает мозг человека «человеческим», обычно отвечают, что в его коре легче устанавливаются связи, чем в коре мозга животного. Такой ответ следует из эмпирической традиции и изучения больших пучков нервных волокон, связывающих между собой разные отделы мозга. Этой распространенной точке зрения противоречат три группы фактов. Перваяз получена в экспериментах на приматах. Эти эксперименты показали, что основные функциональные связи коры головного мозга- это скорее корково-подкорковые, чем кортико-кортикальные связи. Конечно, возможно, что эти результаты относятся только к-мозгу приматов, и если будет обнаружено, что кортико-кортикальные связи являются решающими для психологических процессов… человека, то они будут признаны основным отличием мозга человека.
Однако дальнейший путь исследований будет, вероятно, нестоль прямолинейным. Есть и другие аргументы, которые говорят против гипотезы о функционировании мозга на базе ассоциативных волокон и о нарушении его работы вследствие разъединения этих связей. На протяжении всей этой книги я пытался доказать существование голографических преобразований как средства, при помощи которого нервная система кодирует и перекодирует поступающие сигналы. Одним из свойств голограмм является легкость ассоциативного припоминания. В соответствии этим ассоциации должны иметь место внутри системы, а не между системами, и данные, полученные на обезьянах, подтверждают правильность этой точки зрения (Evarts, 1952; Wegener, 1968). Даже ассоциативные связи между отдельными модальностями больше нарушаются при повреждении первичной проекционной коры, чем при повреждении так называемых ассоциативных зон. Данные нейрофизиологии, полученные на отдельных нейронах, также показывают, что многие клетки в первичных проекционных зонах чувствительны к возбуждениям модальности, отличающейся от основной сенсорной модальности, обслуживаемой этими системами (Spinelli, Starr and Barrett, 1968, рис. XIX-7).
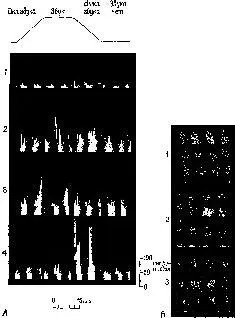
Рис. XIX-7. А – гистограммы ответов четырех различных клеток зрительной коры, чувствительных к изменению звуковых частот. Видно, что клетки 2, 3 и 4 в ответ на модально специфический признак стимула увеличивают частоту разрядов; Б – зрительные рецептивные поля клеток 2, 3 и 4, изображенных на рис. А. Х-ось и У-ось равны 25° зрительного угла. Зрительная ось проходит приблизительно по центру экрана (Spinelli, Start and Barrett, 1968).
Вторая и более прямая линия доказательств получена в результате наблюдений над специфичностью полушарий у человека. Две половины человеческого мозга выполняют разные функции, хотя они и соединяются самым большим пучком ассоциативных волокон мозга – мозолистым телом. Эта мозговая комиссура значительно больше у человека, чем у приматов. Сперри и его сотрудники проделали большие серии экспериментов в целях исследования функций этого нервного тракта путем его перерезки, прерывая таким образом связи между полушариями, которые в результате этого должны работать независимо друг от друга (Sperry, Gazzaniga and Bogen, 1969, рис. XIX-8 и XIX-9).
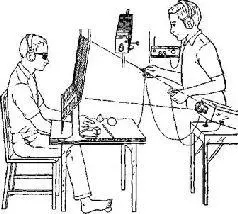
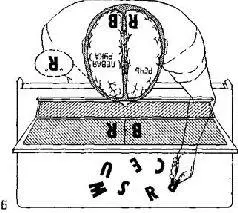
Рис. XIX-8. А – устройство обычной тестирующей -аппаратуры, используемой для демонстрации симптомов комиссуротомии; Б – ручное опознание стимула, подаваемого в левое зрительное поле, успешно осуществляется левой рукой, хотя испытуемый не может назвать объект (Sperry, 1970).
Читать дальшеИнтервал:
Закладка:
Похожие книги на «ЯЗЫКИ МОЗГА Экспериментальные парадоксы и принципы нейропсихологии»
Представляем Вашему вниманию похожие книги на «ЯЗЫКИ МОЗГА Экспериментальные парадоксы и принципы нейропсихологии» списком для выбора. Мы отобрали схожую по названию и смыслу литературу в надежде предоставить читателям больше вариантов отыскать новые, интересные, ещё непрочитанные произведения.











Обсуждение, отзывы о книге «ЯЗЫКИ МОЗГА Экспериментальные парадоксы и принципы нейропсихологии» и просто собственные мнения читателей. Оставьте ваши комментарии, напишите, что Вы думаете о произведении, его смысле или главных героях. Укажите что конкретно понравилось, а что нет, и почему Вы так считаете.