К.ПРИБРАМ - ЯЗЫКИ МОЗГА Экспериментальные парадоксы и принципы нейропсихологии
Здесь есть возможность читать онлайн «К.ПРИБРАМ - ЯЗЫКИ МОЗГА Экспериментальные парадоксы и принципы нейропсихологии» весь текст электронной книги совершенно бесплатно (целиком полную версию без сокращений). В некоторых случаях можно слушать аудио, скачать через торрент в формате fb2 и присутствует краткое содержание. Жанр: Медицина, на русском языке. Описание произведения, (предисловие) а так же отзывы посетителей доступны на портале библиотеки ЛибКат.

- Название:ЯЗЫКИ МОЗГА Экспериментальные парадоксы и принципы нейропсихологии
- Автор:
- Жанр:
- Год:неизвестен
- ISBN:нет данных
- Рейтинг книги:5 / 5. Голосов: 1
-
Избранное:Добавить в избранное
- Отзывы:
-
Ваша оценка:
ЯЗЫКИ МОЗГА Экспериментальные парадоксы и принципы нейропсихологии: краткое содержание, описание и аннотация
Предлагаем к чтению аннотацию, описание, краткое содержание или предисловие (зависит от того, что написал сам автор книги «ЯЗЫКИ МОЗГА Экспериментальные парадоксы и принципы нейропсихологии»). Если вы не нашли необходимую информацию о книге — напишите в комментариях, мы постараемся отыскать её.
Под редакцией и с предисловием
действительного члена АПН СССР
А. Р. Лурия
Издательство «Прогресс» Москва 1975
Редакция литературы по философии
Перевод на русский язык с изменениями. "Прогресс"
ЯЗЫКИ МОЗГА Экспериментальные парадоксы и принципы нейропсихологии — читать онлайн бесплатно полную книгу (весь текст) целиком
Ниже представлен текст книги, разбитый по страницам. Система сохранения места последней прочитанной страницы, позволяет с удобством читать онлайн бесплатно книгу «ЯЗЫКИ МОЗГА Экспериментальные парадоксы и принципы нейропсихологии», без необходимости каждый раз заново искать на чём Вы остановились. Поставьте закладку, и сможете в любой момент перейти на страницу, на которой закончили чтение.
Интервал:
Закладка:
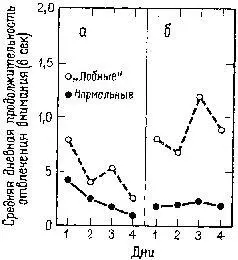
Рис. XVIII-8. График, показывающий среднюю дневную продолжительность отвлечения внимания (средний латентный период отвлечения внимания на пробу минус медиана латентного периода): а – для условий, когда стимул изменяется, а место его остается постоянным; б – для условий, когда изменяется место стимула, а сам ой постоянен (Grueninger and Pribram, 1969).

Рис. XVIII-9. Карты зрительных рецептивных полей, показывающие, как изменяется информация, поступающая по первичным зрительным путям, при стимуляции различных отделов мозга. Карта а – нормальный ответ клетки латерального коленчатого тела при движении источника света по экрану; карта в показывает, как уменьшается рецептивное поле при стимуляции нижневисочной коры; карта б свидетельствует о расширении рецептивного поля, вызванном стимуляцией лобной коры; карта г – конечная контрольная запись, сделанная через 55 мин после записи a (Spinelli and Pribram, 1967).
Приведенный анализ опытов с отсроченным чередованием и отсроченными реакциями позволяет предположить, что повреждение лобных долей вызывает нарушение тех процессов мозга, существенным элементом которых является кодирование изменений состояний. Такие процессы свойственны работе кратковременной памяти, предполагающей скорее решения, зависящие от контекста, чем хорошо упроченные действия, не зависящие от контекста, и нарушения их отражаются как на решении задач, так и на эмоциональном поведении. Можно предположить, что подобный дефект сказывается на решении задач потому, что организм не в состоянии регулировать свое поведение на основе событий, вносящих беспокойство и сигнализирующих об изменениях в контексте. Этот дефект проявляется в эмоциональном поведении: организм не справился с контролем, регистрацией и оценкой изменений, непрерывно усложняющих контекст и таким образом увеличивающих беспокойство в настоящем. При повреждении височной доли утрата способности к восприятию контекста проявляется в том, что поток событий не сохраняется в следах и, таким образом, протекает только в настоящем, которое не имеет ни прошлого, ни будущего. Организм становится автоматом, находящимся во власти своих мгновенных состояний, которые он не может регулировать.
Остается проанализировать связь между кратковременной памятью и символическими процессами. Рассмотрим сначала память. Обычно считают, что процесс сохранения следов происходит примерно так: событие регистрируется в кратковременной памяти, где оно циркулирует в течение некоторого времени, как след в нервных цепях, а затем переводится в более долговременное хранение в молекулярной форме. Этот процесс переноса называется консолидацией следа памяти. Если рассматривать только процессы хранения следов, эту картину можно считать относительно верной. Мы уже отмечали, что привыкание зависит, вероятно, от замкнутой системы с обратной связью, которая уменьшает нервную активность. Ряд таких замкнутых контуров может успешно создать буферный механизм хранения следов в пределах сенсорных систем. Мы также отмечали, что подкрепление может продлить некоторые виды нервной активности, вызывая химические изменения и даже рост глиальной и нервной ткани. Изменения в этих основных механизмах могли бы легко объяснить данные, полученные при экспериментальной проверке гипотезы консолидации.
Но анализ процессов памяти только с точки зрения хранения следов не объясняет другие факты, связанные с запоминанием, – узнавание и припоминание. Узнавание предполагает наложение входного сигнала на след, хранящийся в долговременной памяти. Такое наложение во многих случаях может происходить непосредственно, без вмешательства кратковременной памяти, а это значит, что узнавание осуществляется благодаря параллельной обработке информации механизмом памяти, который может восстанавливать опознанный образ даже на основе информации, лишь частично воспроизводящей хранящийся сигнал. Узнавание, однако, не всегда бывает таким непосредственным. В более сложных ситуациях для узнавания может потребоваться некоторое время и оно может зависеть от более медленных процессов выравнивания кортикальных диполей и других процессов. Этот тип узнавания соответствует большинству классических нейрофизиологических точек зрения на процесс памяти.
Читать дальшеИнтервал:
Закладка:
Похожие книги на «ЯЗЫКИ МОЗГА Экспериментальные парадоксы и принципы нейропсихологии»
Представляем Вашему вниманию похожие книги на «ЯЗЫКИ МОЗГА Экспериментальные парадоксы и принципы нейропсихологии» списком для выбора. Мы отобрали схожую по названию и смыслу литературу в надежде предоставить читателям больше вариантов отыскать новые, интересные, ещё непрочитанные произведения.











Обсуждение, отзывы о книге «ЯЗЫКИ МОЗГА Экспериментальные парадоксы и принципы нейропсихологии» и просто собственные мнения читателей. Оставьте ваши комментарии, напишите, что Вы думаете о произведении, его смысле или главных героях. Укажите что конкретно понравилось, а что нет, и почему Вы так считаете.