К.ПРИБРАМ - ЯЗЫКИ МОЗГА Экспериментальные парадоксы и принципы нейропсихологии
Здесь есть возможность читать онлайн «К.ПРИБРАМ - ЯЗЫКИ МОЗГА Экспериментальные парадоксы и принципы нейропсихологии» весь текст электронной книги совершенно бесплатно (целиком полную версию без сокращений). В некоторых случаях можно слушать аудио, скачать через торрент в формате fb2 и присутствует краткое содержание. Жанр: Медицина, на русском языке. Описание произведения, (предисловие) а так же отзывы посетителей доступны на портале библиотеки ЛибКат.

- Название:ЯЗЫКИ МОЗГА Экспериментальные парадоксы и принципы нейропсихологии
- Автор:
- Жанр:
- Год:неизвестен
- ISBN:нет данных
- Рейтинг книги:5 / 5. Голосов: 1
-
Избранное:Добавить в избранное
- Отзывы:
-
Ваша оценка:
ЯЗЫКИ МОЗГА Экспериментальные парадоксы и принципы нейропсихологии: краткое содержание, описание и аннотация
Предлагаем к чтению аннотацию, описание, краткое содержание или предисловие (зависит от того, что написал сам автор книги «ЯЗЫКИ МОЗГА Экспериментальные парадоксы и принципы нейропсихологии»). Если вы не нашли необходимую информацию о книге — напишите в комментариях, мы постараемся отыскать её.
Под редакцией и с предисловием
действительного члена АПН СССР
А. Р. Лурия
Издательство «Прогресс» Москва 1975
Редакция литературы по философии
Перевод на русский язык с изменениями. "Прогресс"
ЯЗЫКИ МОЗГА Экспериментальные парадоксы и принципы нейропсихологии — читать онлайн бесплатно полную книгу (весь текст) целиком
Ниже представлен текст книги, разбитый по страницам. Система сохранения места последней прочитанной страницы, позволяет с удобством читать онлайн бесплатно книгу «ЯЗЫКИ МОЗГА Экспериментальные парадоксы и принципы нейропсихологии», без необходимости каждый раз заново искать на чём Вы остановились. Поставьте закладку, и сможете в любой момент перейти на страницу, на которой закончили чтение.
Интервал:
Закладка:
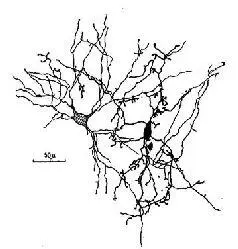
Рис. 1-13. Зарисовка, показывающая многочисленные точки контакта (синаптического и эфаптического) между двумя нейронами мозга (Ralston, 1968).
Таким образом, эфаптические и синаптические процессы, возникающие в местах соединений между нейронами, формируют определенную динамическую структуру, заключение о природе которой может быть сделано на основе знания тонкой структуры мозга и его электрической активности. Можно, в частности, предположить, что эти динамические структуры создают волновые фронты. Это предположение становится особенно полезным при рассмотрении проблем эквивалентности (см. вторую часть книги). Это не значит, что микроструктуру медленного потенциала следует рассматривать в терминах механической волны, но иногда это сходство полезно иметь в виду. Ведь именно интерпретация динамических структур медленных потенциалов как образующих волновые фронты помогает представить себе то, что происходит.
«Нейрон – это реле, действующее по закону «все или ничего». Импульс, достигающий синапса, вызывает очень небольшой и временный электрический эффект равный 0,001 вольта и длящийся от 0,01 до 0,02 сек. Необходимо примерно в 10 раз большее возбуждение, чтобы вызвать разряд Нейрона.
Так как требуется конвергенция многих импульсов на какой-нибудь «дин нейрон, чтобы вызвать его разряд, то цепи, состоящие из одиночных нейронов, не могут обеспечить распространение волны активности в коре.
Распространение активности представляет собой, скорее, передний фронт движения во многих направлениях, когда параллельно активизированы многие клетки в каждом синаптическом звене цепи… (рис. 1-14).

Рис. 1-14. На этой диаграмме синаптические области коры изображены в виде кружков на плоскости. Движение процесса, которое может осуществляться во многих направлениях и характеризуется специфической нейронной структурой возбуждения, показано черными кружками для одного направления и темно-серыми для другого. Области с контурными кружками не активизируются ни одной из динамических структур. Следует отметить, что на пересечении этих двух направлений движения одни и те же области принимали бы участие в обоих процессах и поэтому каждая из них была бы изображена в виде очень большого серого кружка, а не как темно-серый или черный кружок, как показано на рисунке (Eccles, 1970).
Мы сразу же получаем объяснение одного замечательного свойства нервной сети: каким образом два совершенно различных входных воздействия (одно на клетки А, и А 2, другое – на клетки А 3и А 4) могут быть переданы посредством одной и той же динамической структуры клеточных связей, противодействуя друг другу и достигая совершенно различных выходов (Г 3– Г 4) . . . (рис. 1-15).
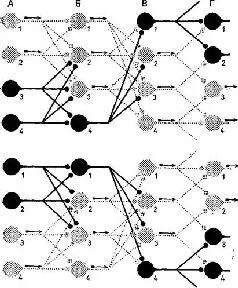
Рис. 1-15. Модель очень схематизированной нейронной сети, иллюстрирующая простейший случай проведения в системе с множеством направлений. На верхней и нижней диаграммах представлены нервные чзети с одной и той же анатомией. В колонках А, Б и В изображены синаптические контакты 12 клеток, клетки генерирующие импульс (со стрелками) – светло-серого цвета, молчащие клетки – черные. Предполагается, что клетка разряжается импульсом, если возбуждается двумя или больше синапсами (также светло-серого цвета). Таким образом, входное воздействие А]А 2 вызывает разряд Г 3 Г 4 (верхняя диаграмма), в то время как вход от А 3 А 4 достигает выхода на Г1Г2 (нижняя диаграмма). Нейроны Бг, Б 3 , Вг, В 3 активизируются в зоне пересечения этих двух входных воздействий. Серьезным недостатком этой диаграммы является то, что она игнорирует тормозные элементы (Eccles, 1968).
Конечно, передача волнового фронта в коре происходит гораздо сложнее. При вовлечении 100 нейронов на каждой релейной станции передняя волна может захватывать более 100 000 нейронов в одну секунду. Такая волна обладает… богатыми потенциальными возможностями…» (Eccles, 4958, р. 4-7).
Другой вывод следует из модели не распространяющейся стоячей волны, основанной на идеализированном представлении об анатомическом распределении связей между нейронами (Scholl, 1956). Все предположения, лежащие в основе этой модели, вполне разумны: 1) нейроны распределены в случайном порядке; 2) богатство связей между клетками уменьшается с расстоянием;. 3) пространственная структура распределения процессов в каждой клетке отличается тем, каким образом она передает возбуждение другим клеткам; 4) наблюдается затухание возбуждения во времени, то есть изменение потенциала в местах соединений имеет конечную длительность, и, наконец, 5) возбуждение само себя поддерживает.
Читать дальшеИнтервал:
Закладка:
Похожие книги на «ЯЗЫКИ МОЗГА Экспериментальные парадоксы и принципы нейропсихологии»
Представляем Вашему вниманию похожие книги на «ЯЗЫКИ МОЗГА Экспериментальные парадоксы и принципы нейропсихологии» списком для выбора. Мы отобрали схожую по названию и смыслу литературу в надежде предоставить читателям больше вариантов отыскать новые, интересные, ещё непрочитанные произведения.











Обсуждение, отзывы о книге «ЯЗЫКИ МОЗГА Экспериментальные парадоксы и принципы нейропсихологии» и просто собственные мнения читателей. Оставьте ваши комментарии, напишите, что Вы думаете о произведении, его смысле или главных героях. Укажите что конкретно понравилось, а что нет, и почему Вы так считаете.