Виктор Бродянский - Вечный двигатель — прежде и теперь. От утопии — к науке, от науки — к утопии
Здесь есть возможность читать онлайн «Виктор Бродянский - Вечный двигатель — прежде и теперь. От утопии — к науке, от науки — к утопии» весь текст электронной книги совершенно бесплатно (целиком полную версию без сокращений). В некоторых случаях можно слушать аудио, скачать через торрент в формате fb2 и присутствует краткое содержание. Город: Москва, Год выпуска: 2001, ISBN: 2001, Издательство: ФИЗМАТЛИТ, Жанр: Физика, на русском языке. Описание произведения, (предисловие) а так же отзывы посетителей доступны на портале библиотеки ЛибКат.

- Название:Вечный двигатель — прежде и теперь. От утопии — к науке, от науки — к утопии
- Автор:
- Издательство:ФИЗМАТЛИТ
- Жанр:
- Год:2001
- Город:Москва
- ISBN:5-9221-0202-8
- Рейтинг книги:4 / 5. Голосов: 1
-
Избранное:Добавить в избранное
- Отзывы:
-
Ваша оценка:
Вечный двигатель — прежде и теперь. От утопии — к науке, от науки — к утопии: краткое содержание, описание и аннотация
Предлагаем к чтению аннотацию, описание, краткое содержание или предисловие (зависит от того, что написал сам автор книги «Вечный двигатель — прежде и теперь. От утопии — к науке, от науки — к утопии»). Если вы не нашли необходимую информацию о книге — напишите в комментариях, мы постараемся отыскать её.
Для широкого круга читателей, интересующихся историей техники и ее современными проблемами.
Вечный двигатель — прежде и теперь. От утопии — к науке, от науки — к утопии — читать онлайн бесплатно полную книгу (весь текст) целиком
Ниже представлен текст книги, разбитый по страницам. Система сохранения места последней прочитанной страницы, позволяет с удобством читать онлайн бесплатно книгу «Вечный двигатель — прежде и теперь. От утопии — к науке, от науки — к утопии», без необходимости каждый раз заново искать на чём Вы остановились. Поставьте закладку, и сможете в любой момент перейти на страницу, на которой закончили чтение.
Интервал:
Закладка:
Вот что пишет по этому поводу в предисловии к книге П.К. Ощепкова [3.1] проф. П. Остроумов: «…Да и среди непосредственно окружающего нас мира мы наблюдаем явления, в которых хаос уступает порядку, где также, хотя и временно, наблюдаются как бы отступления от законов статистики, а теория вероятностей требует углубления и расширения. Это — явления в живой природе. Здесь второй закон в его примитивной форме применим далеко не всегда. Невольно возникает мысль: нельзя ли искусственно создать механизм, упорядочивающий статистическое тепловое движение частиц, воспроизводящий функции живого организма хотя бы лишь с энергетической стороны».
Если опустить весьма неопределенные, но «ученые» слова, не имеющие конкретного содержания, вроде «временного отступления от законов статистики», «примитивной формы второго закона» и «углубления и расширения теории вероятностей», то остается достаточно четкий тезис: живая природа демонстрирует нам антиэнтропийные процессы, противоречащие второму закону; познаем их и сделаем на их основе ppm-2!
Если это так, то нужно внять призывам Остроумова и Ощепкова и развернуть усилия энергетиков в этом многообещающем направлении; если же это не так и живая природа подчиняется второму закону, то следование их призывам бессмысленно и ведет в тупик.
Итак, что же происходит с энтропией в живой природе? Чтобы разобраться в этом, нет никакой необходимости вести специальные исследования: вопрос давно решен и нужно только познакомиться с соответствующей литературой. Наиболее четко существо дела изложено в небольшой, но очень весомой классической книжке известного физика А. Шредингера «Что такое жизнь с точки зрения физика» [1.8]. В 1984 г. вышла в значительной степени посвященная этой же теме научно-популярная работа чл.корр. АН СССР К.К. Ребане [1.10]. Мы подойдем к изложению вопроса не столько с физических, сколько с более конкретных инженерно-термодинамических позиций, имея в виду конечную цель, связанную с ppm-2.
Составим для этого прежде всего в общем виде энергетический баланс, характерный для растений, а затем такой же для животных. Такой баланс можно представить достаточно надежно, если не углубляться в существо сложнейших биологических процессов, а ограничиться входящими и выходящими потоками энергии.
На рис. 4.3 представлена схема материального (потоки вещества) и энергетического балансов растения, основанных на законах сохранения массы и энергии. Чтобы составить такие балансы, окружим растение так называемой замкнутой контрольной поверхностью (штриховая линия), чтобы учесть все входящие и выходящие потоки. Если хотя бы один из них ускользнет от учета (или, наоборот, будет учтен тот, который через контрольную поверхность не проходит), баланс станет неверным. Тогда никаких мало-мальски стоящих выводов из него делать нельзя. Мы постараемся не допустить такой ошибки.
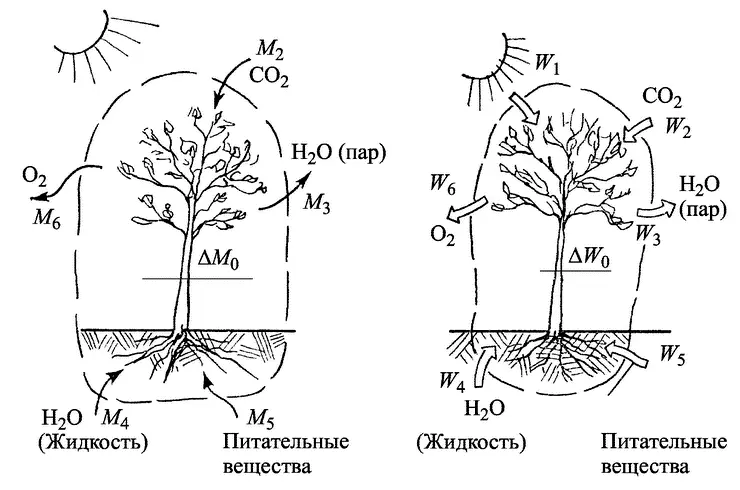
Материальный баланс будет иметь вид
М 2+ М 4+ М 5— (М 3+ М 6) = ΔМ 0.
Это уравнение показывает: все, что получает растение (М 2+ М 4+ М 5) за определенный отрезок времени, за вычетом того, что оно отдает (M 3+ M 6) идет на приращение ΔМ 0его массы, связанное с ростом. Аналогичное уравнение получится и для энергии:
W 1+ W 2+ W 4+ W 5— (W 3+ W 6) = ΔW 0.
Здесь ΔW 0— прирост внутренней энергии растения, определяемый увеличением его массы при росте.
Чтобы установить, нарушает эта система второй закон термодинамики или нет, нужно проверить, что происходит с энтропией в процессе жизнедеятельности растения: увеличивается она или уменьшается?
Очевидно, что живая ткань растения более высоко структурно организована, чем поступающие из воздуха питательные вещества. Поэтому при образовании такой ткани (с массой ΔM 0) ее энтропия будет несомненно меньше, чем суммарная энтропия исходных веществ (СО 2, H 2O) и питательных веществ почвы). В этом смысле образование и накопление живой ткани растения и поддержание ее существования будет, несомненно, антиэнтропийным процессом. Но никак нельзя забывать, что одновременно неизбежно меняется энтропия потоков вещества и энергии, проходящих через контрольную поверхность. Здесь получается обратная картина (рис. 4.3): суммарная энтропия выходящих потоков (3 и 6) неизбежно оказывается много большей, чем входящих (7, 2, 4 и 5). Это объясняется тем, что энтропия поглощаемого солнечного излучения [60] Поток излучения, как и всякий поток энергии, тоже характеризуется определенной степенью беспорядка (разные частоты и другие характеристики колебаний частей спектра). Только монохроматическое когерентное излучение (например, лазера) полностью упорядочено и (как и работа) характеризуется нулевой энтропией.
сравнительно невелика, так же как и поступающих из почвы минеральных солей; энтропии газов — кислорода и СО 2— близки по значениям. Зато энтропия водяного пара, отдаваемого листьями, относительно велика (примерно в 3 раза больше, чем у воды). В результате энтропия потоков, проходящих через контрольную поверхность, возрастает намного больше, чем снижается энтропия веществ, превращающихся в органическую ткань.
Интервал:
Закладка:
Похожие книги на «Вечный двигатель — прежде и теперь. От утопии — к науке, от науки — к утопии»
Представляем Вашему вниманию похожие книги на «Вечный двигатель — прежде и теперь. От утопии — к науке, от науки — к утопии» списком для выбора. Мы отобрали схожую по названию и смыслу литературу в надежде предоставить читателям больше вариантов отыскать новые, интересные, ещё непрочитанные произведения.









Обсуждение, отзывы о книге «Вечный двигатель — прежде и теперь. От утопии — к науке, от науки — к утопии» и просто собственные мнения читателей. Оставьте ваши комментарии, напишите, что Вы думаете о произведении, его смысле или главных героях. Укажите что конкретно понравилось, а что нет, и почему Вы так считаете.