Борис Режабек - Избранные труды. Том 1
Здесь есть возможность читать онлайн «Борис Режабек - Избранные труды. Том 1» — ознакомительный отрывок электронной книги совершенно бесплатно, а после прочтения отрывка купить полную версию. В некоторых случаях можно слушать аудио, скачать через торрент в формате fb2 и присутствует краткое содержание. ISBN: , Жанр: Философия, Прочая научная литература, Биология, на русском языке. Описание произведения, (предисловие) а так же отзывы посетителей доступны на портале библиотеки ЛибКат.

- Название:Избранные труды. Том 1
- Автор:
- Жанр:
- Год:неизвестен
- ISBN:9785005174178
- Рейтинг книги:5 / 5. Голосов: 1
-
Избранное:Добавить в избранное
- Отзывы:
-
Ваша оценка:
Избранные труды. Том 1: краткое содержание, описание и аннотация
Предлагаем к чтению аннотацию, описание, краткое содержание или предисловие (зависит от того, что написал сам автор книги «Избранные труды. Том 1»). Если вы не нашли необходимую информацию о книге — напишите в комментариях, мы постараемся отыскать её.
Избранные труды. Том 1 — читать онлайн ознакомительный отрывок
Ниже представлен текст книги, разбитый по страницам. Система сохранения места последней прочитанной страницы, позволяет с удобством читать онлайн бесплатно книгу «Избранные труды. Том 1», без необходимости каждый раз заново искать на чём Вы остановились. Поставьте закладку, и сможете в любой момент перейти на страницу, на которой закончили чтение.
Интервал:
Закладка:
Хотя этот вопрос несомненно важен, в современных моделях нейронов, к сожалению, не представлены ни способность нейрона к целесообразному приспособлению, ни возможность наличия эндогенных ритмов, свойст- венных данному нейрону и позволяющих ему избирательно реагировать на входной импульсный поток. Наличие таких ритмов обсуждалось в ряде физиологических работ в связи спроблемой «пейсмекеров» [11].
В настоящей работе представлены эксперименталь- ные данные, свидетельствующие о наличии таких ритмов в изолированной нейрорецепторной клетке речного рака.
Методика
Потенциалы действия (ПД) отводились внеклеточно от нервного ствола с помощью серебряного электрода диаметром 0,3 мм. Импульсы усиливались УБП1—02 и регистрировались с помощью шлейфного осциллографа Н-102.
Раздражение производилось стеклянными микроэлек- тродами, заполненными 3 М раствором КСl. Диаметр кончика измерялся под микроскопом МБР-1 и составлял 2—5мк, сопротивление 1—5 Мом.
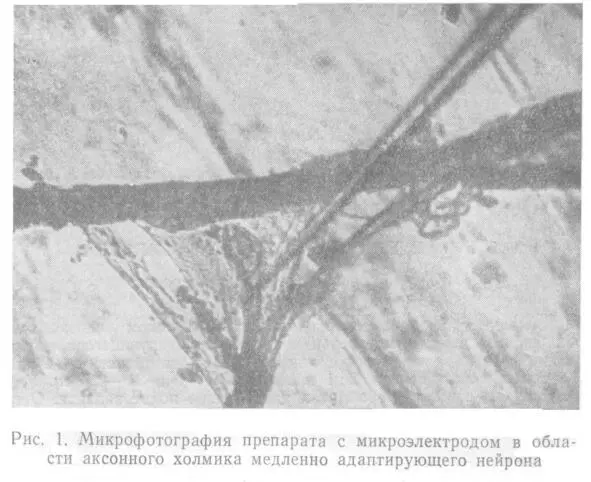
Микроэлектрод подводили с помощью микроманипулятора ММ-1 к избранной точке нейрона под контролем микроскопа МБР-1 (рис.1). В качестве источника раздражающего тока использовался генератор Г3—47.
Результаты экспериментов
– Реакция быстроадаптирующегося нейрона.
Реакция быстроадаптирующегося (БА) нейрона на механичеcкое (адекватное) раздражение достаточно подробно изучена многими авторами [12, 13].
При электрическом раздражении наиболее эффективным является положение микроэлектрода в области аксонного холмика (АХ), при этом возбуждение происходит, когда на микроэлектроде отрицательное напря- жение. Порог раздражения в этом случае минимален. Располагая микроэлектрод вблизи сомы или удаляясь от АХ по волокну, можно наблюдать изменение порога и даже инверсию знака раздражающего потенциала.
При условии одинаковой реакции на толчок постоянного тока, которой можно добиться, расположив микроэлектрод при подготовке к опыту соответствующим образом, реакции различных препаратов на ритмическое раздражение совпадают с точностью до 10%.
Такое раздражение не вызывает импульсной активности нейрона, пока f c <1,5 гц.
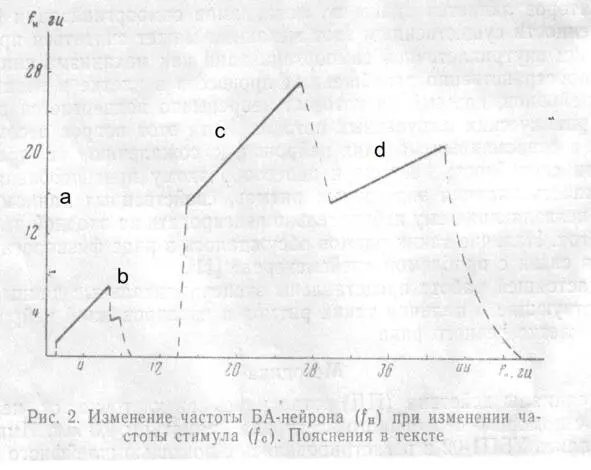
Рис.2
На рис. 2 показано изменение частоты БА-нейрона (f H) при изменении частоты стимула (f c); амплитуда раздражения 11 в. Микроэлектрод расположен вблизи АХ.
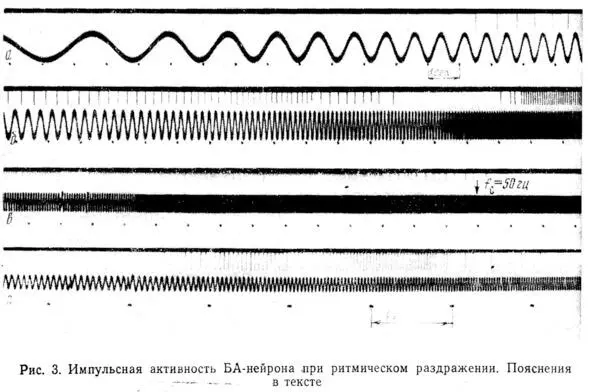
При достижении этой частоты наблюдается резкий переход к режиму, при котором нейрон работает с частотой, совпадающей с f c(рис. 3,а). Диапазон захватывания частоты (ЗЧ) зависит от амплитуды стимула.
В данном опыте ЗЧ скачком прекращается при fc = 7 гц. При изменении f сот 7 до 8 гц наблюдается ЗЧ с делением в отношении 2:1, при частотах, незначительно превышающих 8 гц, нейрон реагирует на каждый третий или четвертый стимул и затем реакция вообще исчезает вплоть до частоты 13 гц (рис. 3,б). От 13 до 14 гц реакция является вероятностной, т. е. однозначного соответствия между f си f Hустановить не удается и при повторных стимуляциях реакция в этом диапазоне оказывается каждый раз иной. С 14 гц начинается ЗЧ вплоть до 27 гц. В диапазоне от 27 до 30 гц наблюдается переходной режим реакции, при котором можно обнаружить отклик нейрона, соответствующий либо каждому периоду стимула, либо с выпадением одного периода. С 30 до 42 гц наблюдается ЗЧ с делением частоты 2:1, и, наконец, при более высоких частотах реакция вновь становится вероятностной.
При дальнейшем увеличении частоты стимуляции вплоть до 20 000 гц импульсная реакция нейрона отсутствует. На рис. 3, в представлен ход процесса от 15 до 60 гц. Если изменять амплитуду стимула, общий ход реакции закономерно изменяется.
Результаты можно объединить с помощью рис.4. Линия а представляет собой геометрическое место точек, соответствующих началу первой полосы ЗЧ с отношением частот f c: fa равным 1:1. Линия б соответ ствует переходу в вероятностный режим, в – началу первой полосы ЗЧ с отношением частот 2:1, г – ее окончанию и д – исчезновению импульсной реакции. Линия е соответствует началу новой полосы ЗЧ.
Читать дальшеИнтервал:
Закладка:
Похожие книги на «Избранные труды. Том 1»
Представляем Вашему вниманию похожие книги на «Избранные труды. Том 1» списком для выбора. Мы отобрали схожую по названию и смыслу литературу в надежде предоставить читателям больше вариантов отыскать новые, интересные, ещё непрочитанные произведения.












Обсуждение, отзывы о книге «Избранные труды. Том 1» и просто собственные мнения читателей. Оставьте ваши комментарии, напишите, что Вы думаете о произведении, его смысле или главных героях. Укажите что конкретно понравилось, а что нет, и почему Вы так считаете.