Максим Франк-Каменецкий - Самая главная молекула. От структуры ДНК к биомедицине XXI века
Здесь есть возможность читать онлайн «Максим Франк-Каменецкий - Самая главная молекула. От структуры ДНК к биомедицине XXI века» — ознакомительный отрывок электронной книги совершенно бесплатно, а после прочтения отрывка купить полную версию. В некоторых случаях можно слушать аудио, скачать через торрент в формате fb2 и присутствует краткое содержание. Город: Москва, Год выпуска: 2017, ISBN: 2017, Издательство: Литагент Альпина, Жанр: Биология, Биология, Прочая научная литература, на русском языке. Описание произведения, (предисловие) а так же отзывы посетителей доступны на портале библиотеки ЛибКат.

- Название:Самая главная молекула. От структуры ДНК к биомедицине XXI века
- Автор:
- Издательство:Литагент Альпина
- Жанр:
- Год:2017
- Город:Москва
- ISBN:978-5-9614-4522-0
- Рейтинг книги:3 / 5. Голосов: 1
-
Избранное:Добавить в избранное
- Отзывы:
-
Ваша оценка:
Самая главная молекула. От структуры ДНК к биомедицине XXI века: краткое содержание, описание и аннотация
Предлагаем к чтению аннотацию, описание, краткое содержание или предисловие (зависит от того, что написал сам автор книги «Самая главная молекула. От структуры ДНК к биомедицине XXI века»). Если вы не нашли необходимую информацию о книге — напишите в комментариях, мы постараемся отыскать её.
Самая главная молекула. От структуры ДНК к биомедицине XXI века — читать онлайн ознакомительный отрывок
Ниже представлен текст книги, разбитый по страницам. Система сохранения места последней прочитанной страницы, позволяет с удобством читать онлайн бесплатно книгу «Самая главная молекула. От структуры ДНК к биомедицине XXI века», без необходимости каждый раз заново искать на чём Вы остановились. Поставьте закладку, и сможете в любой момент перейти на страницу, на которой закончили чтение.
Интервал:
Закладка:
В 2009 году Рогаеву удалось точно выяснить, в чем заключалась зловредная мутация. Это была замена одного нуклеотида (А заменен на Г) в третьем с конца положении в интроне, находящемся между экзонами #3 и #4, гена, кодирующего фактор свертываемости крови IX (рис. 49). В результате этой замены возникает новый сигнал сплайсинга на конце интрона, и сплайсинг происходит неверно: в результирующей молекуле мРНК между экзонами #3 и #4 вклиниваются два нуклеотида. Это приводит к тому, что сразу после экзона #3 сбивается рамка считывания мРНК рибосомой, в белковую цепь включаются совсем другие аминокислоты, а вскоре вообще появляется терминирующий кодон, и белковая цепь обрывается (рис. 49). Получается не просто мутантный белок, а вообще нечто не имеющее ничего общего с нормальным белком. Вот такой зловреднейшей мутацией наградил свою дочь граф Кентский Эдвард в 1818 году. И что ему взбрендило за год до своей смерти ребенка заводить, честное слово?!
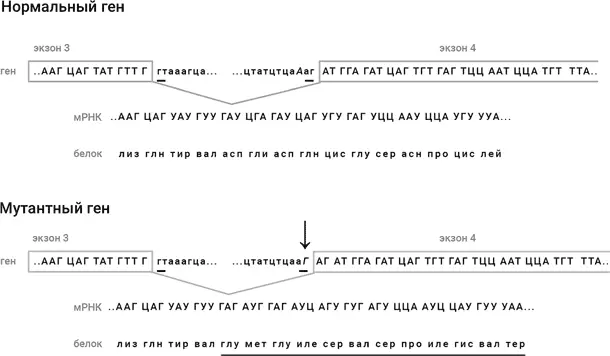
Рис. 49.Мутация в геноме королевы Виктории, приведшая к тому, что царевич Алексей оказался болен гемофилией. Сверху показана последовательность нормального гена фактора свертываемости крови IX, а внизу – последовательность мутантного гена. Последовательности отличаются только одной буквой, которая помечена стрелкой и дана курсивом: буква А в нормальном гене заменена в мутантном гене на Г. Последовательность нуклеотидов в интроне, расположенном между экзонами #3 и #4, дана строчными буквами. Подчеркнутые крайние пары букв в интроне, гт и аг, служат сигналами сплайсинга. Мутация приводит к тому, что возникает новый сигнал сплайсинга аГ на две буквы раньше правильного сигнала. В результате в самом начале экзона #4 вклиниваются два нуклеотида из интрона: АГ. Поэтому после места сплайсинга экзонов #3 и #4 сбивается рамка считывания мРНК рибосомой, и синтезируется аминокислотная последовательность, совершенно отличная от последовательности нормального фактора свертываемости крови (подчеркнуто), а потом вообще появляется терминирующий кодон УАА, и белковая цепь обрывается
Еврейские гены
Открытие двойной спирали Уотсоном и Криком было самым главным, но не единственным прорывом в нашем понимании феномена жизни. Задолго до этого, в 60-х годах XIX века, возникли две теории – Чарльза Дарвина и Грегора Менделя, которые привели к пониманию того, что необыкновенное разнообразие форм жизни является следствием эволюционного процесса. Краеугольным камнем эволюционной теории являются концепции изменчивости и отбора. Изменчивость происходит благодаря мутациям в генах, которые представляют собой, как мы теперь знаем, изменения последовательности нуклеотидов в ДНК, а затем отбор приводит к выживанию наиболее приспособленных мутантов. Успехи эволюционной теории в объяснении происхождения видов привели к тому, что концепция отбора наиболее приспособленных стала применяться для объяснения не только происхождения видов, но и для понимания распространенности тех или иных генов внутри человеческих популяций. Хотя описано множество генетических болезней, ярких примеров таких болезней с большим количеством случаев известно не так уж много. Долгое время наиболее известным примером была, да и остается серповидно-клеточная анемия (СКА), и факт ее широкого распространения среди населения Северной Африки (и их потомков афроамериканцев) многие десятилетия служил убедительным аргументом в пользу того, что принцип отбора наиболее приспособленных прекрасно работает в случае генетических болезней. Дело в том, что ареал распространения СКА в Африке с высокой точностью совпадает с ареалом распространения малярии.
В главе 2 мы уже обсуждали мутацию, приводящую к СКА. В отличие от гена гемофилии, ген СКА расположен не в половой хромосоме, а в одной из аутосом (так называют все хромосомы, кроме половых хромосом). У аутосомного гена всегда есть двойник, другой аллельный вариант, как говорят генетики. Если оба аллеля несут СКА-мутацию, т. е. имеет место гомозиготная ситуация, то такой человек заболевает СКА, и его дни сочтены. В случае же гетерозиготной ситуации, когда только один из двух аллелей несет мутацию, не только нормального гемоглобина оказывается достаточно для переноса кислорода, но наличие мутантного гемоглобина каким-то образом защищает от заражения малярией. Механизм защиты не вполне понятен, но сам факт надежно установлен. Так что накопление в популяции гетерозигот по гену СКА является чисто дарвиновским ответом на давление отбора со стороны малярии. Ну а неизбежное появление летальных гомозигот – это вполне приемлемая с точки зрения популяции в целом плата за защиту гетерозигот от малярии.
Читать дальшеИнтервал:
Закладка:
Похожие книги на «Самая главная молекула. От структуры ДНК к биомедицине XXI века»
Представляем Вашему вниманию похожие книги на «Самая главная молекула. От структуры ДНК к биомедицине XXI века» списком для выбора. Мы отобрали схожую по названию и смыслу литературу в надежде предоставить читателям больше вариантов отыскать новые, интересные, ещё непрочитанные произведения.












Обсуждение, отзывы о книге «Самая главная молекула. От структуры ДНК к биомедицине XXI века» и просто собственные мнения читателей. Оставьте ваши комментарии, напишите, что Вы думаете о произведении, его смысле или главных героях. Укажите что конкретно понравилось, а что нет, и почему Вы так считаете.