М. Канунго - Биохимия старения
Здесь есть возможность читать онлайн «М. Канунго - Биохимия старения» весь текст электронной книги совершенно бесплатно (целиком полную версию без сокращений). В некоторых случаях можно слушать аудио, скачать через торрент в формате fb2 и присутствует краткое содержание. Город: Москва, Год выпуска: 1982, Жанр: Биология, Медицина, на русском языке. Описание произведения, (предисловие) а так же отзывы посетителей доступны на портале библиотеки ЛибКат.

- Название:Биохимия старения
- Автор:
- Жанр:
- Год:1982
- Город:Москва
- ISBN:нет данных
- Рейтинг книги:5 / 5. Голосов: 1
-
Избранное:Добавить в избранное
- Отзывы:
-
Ваша оценка:
Биохимия старения: краткое содержание, описание и аннотация
Предлагаем к чтению аннотацию, описание, краткое содержание или предисловие (зависит от того, что написал сам автор книги «Биохимия старения»). Если вы не нашли необходимую информацию о книге — напишите в комментариях, мы постараемся отыскать её.
Предназначена для биологов, биохимиков, геронтологов, врачей-гериатров.
Биохимия старения — читать онлайн бесплатно полную книгу (весь текст) целиком
Ниже представлен текст книги, разбитый по страницам. Система сохранения места последней прочитанной страницы, позволяет с удобством читать онлайн бесплатно книгу «Биохимия старения», без необходимости каждый раз заново искать на чём Вы остановились. Поставьте закладку, и сможете в любой момент перейти на страницу, на которой закончили чтение.
Интервал:
Закладка:
В растворе гистоны разных типов связываются попарно [94], причем наиболее сильная связь наблюдается между гистонами Н3 и Н4. Нуклеосома имеет ось симметрии второго порядка. В детальных рентгеноструктурных и электронно-микроскопических исследованиях кристаллических препаратов нуклеосом [117] показано, что сердцевина нуклеосомы представляет собой плоский клинообразный диск размером 5,7×11×11 нм. Полагают [117], что 140 пар оснований ДНК составляют 1,75 витка спирали, диаметр витка равен 9 нм, а его шаг — 2,8 нм (рис. 2.3). Это соответствует приблизительно 80 парам оснований на сверхспиральный виток В-формы ДНК. Гистоны частично погружены в большую бороздку ДНК, а малая бороздка остается открытой. Брем [50] считает, что сердцевина нуклеосомы имеет клинообразную форму, ее размеры 5,5×10×12 нм, а 140 пар оснований ДНК расположены в виде витка. Длина 140 пар оснований ДНК в 6–7 раз превышает размеры нуклеосомной сердцевины. Таким образом, ДНК конденсирована в 6–7 раз, что обусловлено ее связыванием с основными участками цепей восьми молекул гистонов и закручиванием вокруг сердцевины сверхспирали [337]. Это обеспечивает защиту нуклеосомной ДНК от микрококковой и стафилококковой ДНКаз. Однако панкреатическая ДНКаза I расщепляет эту ДНК с образованием фрагментов, состоящих из десяти нуклеотидов. Из-за спиральной структуры ДНК разные участки ее цепи отличаются друг от друга по чувствительности к ДНКазе I [238]. По-видимому, внутри нуклеосомы имеются отдельные центры, по которым происходит расщепление под действием ДНКазы. ДНКаза II расщепляет нуклеосомную ДНК с образованием двух фрагментов по 100 пар оснований [15]. С помощью гидродинамических методов показано [135], что нуклеосома претерпевает два конформационных перехода, зависящих от концентрации соли. Это служит дополнительным доказательством того, что нуклеосома включает две субчастицы, или половины.
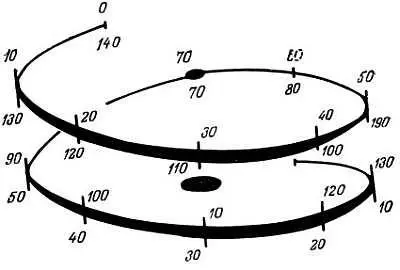
Рис. 2.3. Предполагаемое закручивание суперспирали ДНК вокруг сердцевины нуклеосомы. Отмечены места расщепления ДНК нуклеазой [117]
Сердцевина нуклеосомы содержит по две молекулы каждого из Н2А-, Н2В-, Н3- и Н4-гистонов, которые образуют октамер. Положительно заряженные вытянутые цепи этих гистонов электростатически связаны с отрицательно заряженной ДНК. Полагают, что четыре гистона расположены относительно ДНК следующим образом:
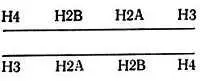
Два гистона, Н3 и Н4, богатые аргинином, вероятно, взаимодействуют с двумя концами фрагмента ДНК. Когда эти гистоны добавляют к двухцепочечной ДНК, они образуют характерную структуру типа бублика, видимую в электронный микроскоп [129]. При воссоединении гистонов сердцевины со 140 парами оснований ДНК образуются частицы, имеющие тот же самый коэффициент седиментации, что и нуклеосомы, полученные из хроматина [36, 345]. Было также показано, что одни гистоны Н3 и Н4 образуют с ДНК структуры, похожие на сердцевины нуклеосом, устойчивые к трипсину [64, 327] и дающие картину дифракции рентгеновских лучей, похожую на картину для нативных нуклеосом [261]. Когда гистоны Н3 и Н4 добавляют к ДНК, они связываются со 140 парами оснований ДНК, которая имеет 1,5 сверхспиральных оборота вокруг тетрамера [195]. Образующаяся структура представляет собой цилиндр с размерами 45×8×8 нм. При последующем добавлении гистонов Н2А и Н2В цилиндр сжимается и становится похожим на нативную нуклеосому. Аналогичные явления наблюдал Картер [70]. Это согласуется с высказанным ранее [198] предположением, что гистоны Н3 и Н4 играют существенную роль в образовании структуры нуклеосомы. Эти два гистона наиболее консервативны, содержат большое количество β-структур и взаимодействуют друг с другом сильнее, чем с другими гистонами. По степени связывания с ДНК гистоны располагаются в следующем порядке: Н3 и Н4>Н2А>Н2В>Н1 [283]. При изучении поперечных сшивок показано, что связаны следующие пары: Н3-Н4, Н2А-Н2В и Н2В-Н4 [84].
Согласно одной из точек зрения, сначала 2 молекулы гистона Н3 и 2 молекулы гистона Н4 образуют тетрамер и связываются со 140 парами оснований ДНК, формируя основную сердцевину нуклеосомы. На втором этапе в эту структуру включаются по две молекулы гистонов Н2А и Н2В, чем и завершается образование нуклеосомы [42, 64, 258, 372]. При изучении сборки новореплицированного хроматина Drosophila показано, что гистоны Н3 и Н4 соединяются с ДНК в течение или вскоре после ее синтеза, гистоны Н2А и Н2В — на 2-10 мин позже, а гистон Н1 — через 10–20 мин, и в результате образуется зрелый хроматин [375]. По-видимому, во взаимодействие с ДНК вовлечены COOH-концы четырех гистонов, так как удаление ЫН2-концевых участков цепей гистонов не влияет на структуру нуклеосомы [371]. Гистоны Н2А и Н2В образуют димеры, взаимодействуя своими центральными неполярными областями, так что NH 2- и COOH-концы остаются свободными. Гистоны Н3 и Н4 образуют димеры путем образования связей между их центральными неполярными областями и COOH-концами, так что основные NH 2-концевые области нуклеосомных гистонов доступны для взаимодействия с кислотными группами ДНК [72]. Роль NH 2-концевых областей четырех гистонов пока не установлена, хотя известно, что они связываются с ДНК. Мирзабеков и др. [252] путем ковалентных сшивок гистонов с 5′-концевыми фрагментами ДНК показали, что каждый гистон связан с 10 парами оснований ДНК. Сборка нуклеосом, по-видимому, контролируется НГБ. Так, очищенный препарат этих белков, выделенный из яиц Xenopus laevis , в бесклеточной системе в присутствии гистонов и очищенной ДНК катализирует образование нуклеосом [217].
Читать дальшеИнтервал:
Закладка:
Похожие книги на «Биохимия старения»
Представляем Вашему вниманию похожие книги на «Биохимия старения» списком для выбора. Мы отобрали схожую по названию и смыслу литературу в надежде предоставить читателям больше вариантов отыскать новые, интересные, ещё непрочитанные произведения.





![Ольга Шестова - Красивое долголетие [10С против старения] [litres]](/books/407424/olga-shestova-krasivoe-dolgoletie-10s-protiv-star-thumb.webp)






Обсуждение, отзывы о книге «Биохимия старения» и просто собственные мнения читателей. Оставьте ваши комментарии, напишите, что Вы думаете о произведении, его смысле или главных героях. Укажите что конкретно понравилось, а что нет, и почему Вы так считаете.