Сергей Ястребов - От атомов к древу. Введение в современную науку о жизни
Здесь есть возможность читать онлайн «Сергей Ястребов - От атомов к древу. Введение в современную науку о жизни» — ознакомительный отрывок электронной книги совершенно бесплатно, а после прочтения отрывка купить полную версию. В некоторых случаях можно слушать аудио, скачать через торрент в формате fb2 и присутствует краткое содержание. Город: Москва, Год выпуска: 2018, ISBN: 2018, Издательство: Альпина нон-фикшн, Жанр: Биология, sci_biochem, sci_popular, на русском языке. Описание произведения, (предисловие) а так же отзывы посетителей доступны на портале библиотеки ЛибКат.

- Название:От атомов к древу. Введение в современную науку о жизни
- Автор:
- Издательство:Альпина нон-фикшн
- Жанр:
- Год:2018
- Город:Москва
- ISBN:978-5-9614-5286-0
- Рейтинг книги:5 / 5. Голосов: 1
-
Избранное:Добавить в избранное
- Отзывы:
-
Ваша оценка:
От атомов к древу. Введение в современную науку о жизни: краткое содержание, описание и аннотация
Предлагаем к чтению аннотацию, описание, краткое содержание или предисловие (зависит от того, что написал сам автор книги «От атомов к древу. Введение в современную науку о жизни»). Если вы не нашли необходимую информацию о книге — напишите в комментариях, мы постараемся отыскать её.
Рекомендуется широкому кругу читателей, всерьез интересующихся современной биологией.
От атомов к древу. Введение в современную науку о жизни — читать онлайн ознакомительный отрывок
Ниже представлен текст книги, разбитый по страницам. Система сохранения места последней прочитанной страницы, позволяет с удобством читать онлайн бесплатно книгу «От атомов к древу. Введение в современную науку о жизни», без необходимости каждый раз заново искать на чём Вы остановились. Поставьте закладку, и сможете в любой момент перейти на страницу, на которой закончили чтение.
Интервал:
Закладка:
Обратившись к литературе, мы сразу увидим, что это отнюдь не фантазия на пустом месте. В последнее десятилетие XX века подобные гипотезы были удивительно популярны, да и сейчас у них есть осторожные сторонники [259] Sogin M. L. Early evolution and the origin of eukaryotes // Current Opinion in Genetics & Development, 1991, V. 1, № 4, 457–463.
[260] Gupta R. S. et al. Cloning of Giardia lamblia heat shock protein HSP70 homologs: implications regarding origin of eukaryotic cells and of endoplasmic reticulum // Proceedings of the National Academy of Sciences , 1994, V. 91, № 8, 2895–2899.
[261] Lake J. A., Rivera M. C. Was the nucleus the first endosymbiont? // Proceedings of the National Academy of Sciences , 1994, V. 91, № 8, 2880–2881.
[262] Moreira D., Lopez-Garcia P. Symbiosis between methanogenic archaea and δ-proteobacteria as the origin of eukaryotes: the syntrophic hypothesis // Journal of Molecular Evolution , 1998, V. 47, № 5, 517–530.
[263] Lopez-Garcia P., Moreira D. Metabolic symbiosis at the origin of eukaryotes // Trends in Biochemical Sciences , 1999, V. 24, № 3, 88–93.
[264] Lake J. A. Eukaryotic origins // Philosophical Transactions of the Royal Society of London, B: Biological Sciences , 2015, V. 370, № 1678, 20140321.
[265] Lopez-Garcia P., Moreira D. Open questions on the origin of eukaryotes // Trends in Ecology & Evolution , 2015, V. 30, № 11, 697–708.
. Существует целое семейство гипотез, согласно которым клеточное ядро имеет симбиотическое происхождение — от археи, встроившейся в клетку более крупной бактерии и окруженной ее сомкнутыми выростами.
Образцом такой гипотезы можно считать эволюционный сценарий, который опубликовали в 1996 году Радхей Гупта и помогавший ему Брайан Голдинг; у этих авторов есть полная аргументация и нет ничего лишнего [266] Gupta R. S., Golding G. B. The origin of the eukaryotic cell // Trends in Biochemical Sciences , 1996, V. 21, № 5, 166–171.
. Гупта и Голдинг изучали эволюцию белков, сравнивая их аминокислотные последовательности. И они пришли к выводу, который в основном подтверждается и сегодня: примерно половина эукариотных белков восходит к археям, а другая половина — к грамотрицательным бактериям. Причем это соотношение настолько близко к равному, что классическая теория (согласно которой архейный предок эукариот всего лишь захватил бактериального симбионта как некий «предмет роскоши») становится неубедительной.
Кроме того, это соотношение трудно объяснить простым горизонтальным переносом генов (даже при том, что интенсивность этого процесса могла быть в древние времена намного выше, чем сейчас). Такой перенос шел бы более-менее равномерно между всеми участниками сообщества. И если бы дело было только в нем, то предок эукариот «нахватался» бы генов от самых разных соседей-бактерий — и грамположительных, и грамотрицательных. Между тем Гупта и Голдинг обнаружили, что у эукариот очень много белков и генов, каким-то образом унаследованных от грамотрицательных бактерий, а вот от грамположительных не нашлось на тот момент ни одного. Конечно, эта избирательность не случайна, вопрос только в ее источнике.
В общем, состав эукариотного генома дает все основания полагать, что при возникновении эукариот клетки археи и грамотрицательной бактерии просто слились (см. рис. 14.4). Получилась единая клетка, сначала унаследовавшая геномы обоих «родителей» — Гупта и Голдинг прямо так их и называют. Но, поскольку два полнофункциональных генома клетке были не нужны, часть каждого из них исчезла, а между оставшимися частями произошло разделение функций. От архейного генома в эукариотной клетке остались в основном «информационные» гены, обеспечивающие работу самого генетического аппарата. А от бактериального генома — в основном «операционные» гены, обеспечивающие обмен веществ. Тогда будет вполне разумно предположить, что в химерной структуре (каковой эукариотная клетка в любом случае несомненно является) от археи произошло ядро, а от бактерии — цитоплазма вместе с наружной мембраной. И получается, что это именно архея проникла внутрь бактерии. Вероятно, сначала она жила во впячивании наружной мембраны бактерии — этаком «заливе». Потом глубокие складки мембраны бактерии, со всех сторон охватившие архею, сомкнулись друг с другом, образовав внутреннюю мембранную систему — ядро и эндоплазматическую сеть. После этого плазматическая мембрана самой археи стала лишней и исчезла. Фактически от археи остался голый генетический аппарат, заключенный внутри системы бактериальных мембран. Ну а гены самой бактерии, сначала находившиеся снаружи от ядра, постепенно мигрировали в него (молекулярные механизмы, делающие возможной такую миграцию, известны). И получилась нормальная эукариотная клетка.
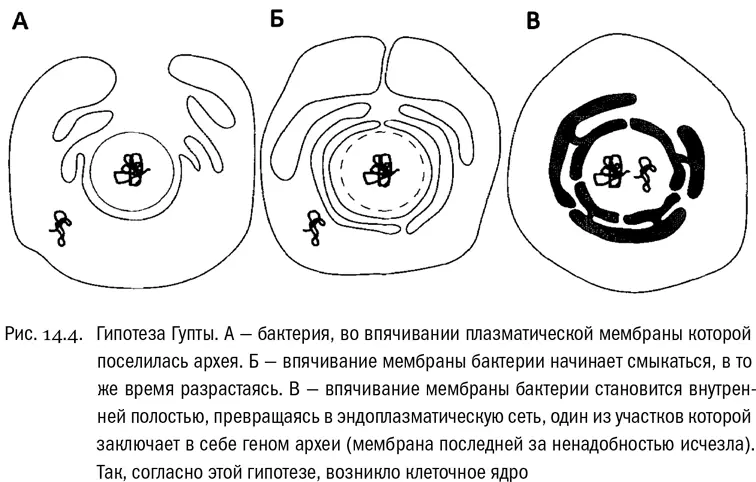
Интересно, что в гипотезе Гупты (назовем ее так для краткости, хотя ее поддерживали и другие ученые) есть кое-что общее с уже знакомой нам гипотезой Баумов. И там и там получается, что остаток исходной архейной клетки находится в основном в эукариотном ядре. Хотя приводящие к этому выводу сценарии прямо-таки полярно противоположны. Гипотеза Баумов, при всем ее бесспорном изяществе, по части состава мембран (архейные vs. бактериальные) дает предсказания, строго обратные тому, что наблюдается в действительности, и выйти из этого положения она может только путем добавления «эпициклов».
Читать дальшеИнтервал:
Закладка:
Похожие книги на «От атомов к древу. Введение в современную науку о жизни»
Представляем Вашему вниманию похожие книги на «От атомов к древу. Введение в современную науку о жизни» списком для выбора. Мы отобрали схожую по названию и смыслу литературу в надежде предоставить читателям больше вариантов отыскать новые, интересные, ещё непрочитанные произведения.










Обсуждение, отзывы о книге «От атомов к древу. Введение в современную науку о жизни» и просто собственные мнения читателей. Оставьте ваши комментарии, напишите, что Вы думаете о произведении, его смысле или главных героях. Укажите что конкретно понравилось, а что нет, и почему Вы так считаете.