Чарльз Эллис - Эпигенетика
Здесь есть возможность читать онлайн «Чарльз Эллис - Эпигенетика» весь текст электронной книги совершенно бесплатно (целиком полную версию без сокращений). В некоторых случаях можно слушать аудио, скачать через торрент в формате fb2 и присутствует краткое содержание. Город: Москва, Год выпуска: 2010, ISBN: 2010, Издательство: Техносфера, Жанр: Биология, sci_popular, на русском языке. Описание произведения, (предисловие) а так же отзывы посетителей доступны на портале библиотеки ЛибКат.
- Название:Эпигенетика
- Автор:
- Издательство:Техносфера
- Жанр:
- Год:2010
- Город:Москва
- ISBN:978-5-94836-257-1
- Рейтинг книги:4 / 5. Голосов: 1
-
Избранное:Добавить в избранное
- Отзывы:
-
Ваша оценка:
Эпигенетика: краткое содержание, описание и аннотация
Предлагаем к чтению аннотацию, описание, краткое содержание или предисловие (зависит от того, что написал сам автор книги «Эпигенетика»). Если вы не нашли необходимую информацию о книге — напишите в комментариях, мы постараемся отыскать её.
Книга ярко и наглядно повествует о новой науке общебиологического значения — эпигенетике, а также об ее отдельных областях. В издании представлено описание разных эпигенетических сигналов и механизмов их реализации, а также собственно феномен, история и концепции эпигенетики, ее отдельные механизмы и пути реализации эпигенетических сигналов в клетке. Авторы различных глав данной книги — ведущие в мире специалисты в области эпигенетики, являющиеся, как правило, и основоположниками ее отдельных областей.
Издание будет полезно широкому кругу читателей, интересующихся коренными проблемами живого мира, сущности жизни и молекулярных механизмов ее проявления.
По формирующейся традиции современной российской научной литературы, оригинальное русскоязычное печатное издание неопрятно переведено, отвратительно вычитано и содержит большое количество ошибок, начиная с обложки. Чарльз Дэвид Эллис указан как С. Д. Эллис.
Эпигенетика — читать онлайн бесплатно полную книгу (весь текст) целиком
Ниже представлен текст книги, разбитый по страницам. Система сохранения места последней прочитанной страницы, позволяет с удобством читать онлайн бесплатно книгу «Эпигенетика», без необходимости каждый раз заново искать на чём Вы остановились. Поставьте закладку, и сможете в любой момент перейти на страницу, на которой закончили чтение.
Интервал:
Закладка:
Оставалось неясным, каким образом, с использованием этих механизмов, информация о состоянии активности могла бы передаваться при клеточном делении; была, таким образом, неясна их роль в эпигенетической передаче информации. Следующим важным шагом стало понимание того, что модифицированные гистоны рекрутируют специфичным в отношении данной модификации образом белки, которые могут влиять на локальные структурные и функциональные состояния хроматина. Было, например, обнаружено, что метилирование лизина 9 в гистоне H3 приводит к рекрутированию белка НР1 гетерохроматина (Bannister et al., 2001; Lachner et al., 2001; Nakayama et al., 2001). Более того, HP1 мог рекрутировать энзим (Suv39hl), отвечающий за метилирование. Это привело к созданию модели воспроизведения сайленсированного состояния хроматина по всему данному участку посредством процессивного [processive] механизма (рис. 2.1а). В равной степени важно, что это обеспечило разумное объяснение того, каким образом это состояние может быть передано и сохранено в цикле репликации (рис. 2.16). Были предложены аналогичные механизмы для воспроизведения активного состояния, включающие метилирование лизина 4 в гистоне H3 и рекрутирование белков группы Trithorax (Wysocka et al., 2005).
Были предложены различные типы механизмов воспроизведения, которые зависели не от модифицированных, а от вариантных гистонов (Ahmad and Henikoff, 2002; McKittrick et al., 2004). Гистон H3 включается в хроматин только во время репликации ДНК. Напротив, вариант этого гистона, H3.3, отличающийся от H3 четырьмя аминокислотами, включается в нуклеосомы не зависящим от репликации образом и имеет тенденцию накапливаться в активном хроматине, где он обогащается «активными» модификациями гистонов (McKittrick et al., 2004). Предположили, что для поддержания активного состояния достаточно присутствия H3.3 и что после репликации остается достаточно H3.3 для поддержания активного состояния, хотя он и разбавляется вдвое. Последующая транскрипция приводила бы к замещению нуклеосом, содержащих H3, вариантом H3.3, воспроизводя таким образом это активное состояние в следующем поколении.
6. Все механизмы взаимосвязаны
В этих моделях в конечном счете начала замыкаться связь между модифицированными или вариантными гистонами, активацией специфических генов и эпигенетикой, хотя, конечно, многое еще остается сделать. В то время как эти механизмы дают нам какие-то представления о том, как состояние гетерохроматина может поддерживаться, они не объясняют, каким образом структуры «молчащего» хроматина устанавливаются впервые. Лишь недавно стало ясно, что это связано с продукцией РНК-транскриптов, особенно с повторяющихся последовательностей, которые подвергаются процессингу в малые РНК благодаря действию таких белков, как Dicer, Argonaute и РНК-зависимая РНК-полимераза. Впоследствии эти РНК рекрутируются в гомологичные сайты ДНК как часть комплексов, включающих компоненты группы белков Polycomb, инициируя таким образом формирование гетерохроматина. Сейчас имеются также данные о том, что для поддержания по крайней мере некоторых гетерохроматиновых районов требуются те же механизмы. В известном смысле эти устойчивые циклические цепи реакций напоминают предложенную Дельбрюком модель 50-летней давности — модель устойчивого биохимического цикла, поддерживающего данное состояние организма.
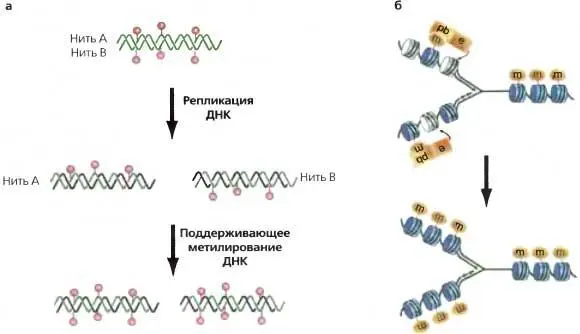
Рис. 2.1.Механизмы поддержания паттерна метилирования ДНК и модификаций гистонов в ходе репликации ДНК
(а) механизм поддержания паттерна метилирования ДНК в ходе репликации ДНК. Во время репликации отдельные нити ДНК, со специфическим паттерном метилирования в остатках CpG или CpXpG, спариваются с нитью вновь синтезированной, неметилированной ДНК. CpG на одной нити имеет соответствующий CpG на другой. Поддерживающая ДНК-метилтрансфераза распознает полуметилированный сайт и метилирует цитозин на новой нити, так что паттерн метилирования не нарушается; (б) общий механизм для поддержания модификаций гистонов в ходе репликации. Модифицированный гистоновый «хвост» (m) взаимодействует с белком-связкой (pb — a protein binder), имеющим сайт связывания, специфичный для данной модификации, pb. в свою очередь, имеет специфический сайт для энзима (е), который выполняет эту модификацию гистона, т. е. в свою очередь, может затем модифицировать соседнюю нуклеосому. Во время репликации вновь откладывающиеся гистоны, которые перемежаются с родительскими гистонами, могут таким образом приобретать данную родительскую модификацию. Сходный механизм мог бы обеспечить воспроизведение, распространение модификаций гистонов с модифицированного на немодифицированный участок на любой стадии клеточного цикла
Читать дальшеИнтервал:
Закладка:
Похожие книги на «Эпигенетика»
Представляем Вашему вниманию похожие книги на «Эпигенетика» списком для выбора. Мы отобрали схожую по названию и смыслу литературу в надежде предоставить читателям больше вариантов отыскать новые, интересные, ещё непрочитанные произведения.











Обсуждение, отзывы о книге «Эпигенетика» и просто собственные мнения читателей. Оставьте ваши комментарии, напишите, что Вы думаете о произведении, его смысле или главных героях. Укажите что конкретно понравилось, а что нет, и почему Вы так считаете.