Кирилл Еськов - Доказательства эволюции
Здесь есть возможность читать онлайн «Кирилл Еськов - Доказательства эволюции» весь текст электронной книги совершенно бесплатно (целиком полную версию без сокращений). В некоторых случаях можно слушать аудио, скачать через торрент в формате fb2 и присутствует краткое содержание. Год выпуска: 2010, Жанр: Биология, на русском языке. Описание произведения, (предисловие) а так же отзывы посетителей доступны на портале библиотеки ЛибКат.

- Название:Доказательства эволюции
- Автор:
- Жанр:
- Год:2010
- ISBN:нет данных
- Рейтинг книги:4 / 5. Голосов: 1
-
Избранное:Добавить в избранное
- Отзывы:
-
Ваша оценка:
Доказательства эволюции: краткое содержание, описание и аннотация
Предлагаем к чтению аннотацию, описание, краткое содержание или предисловие (зависит от того, что написал сам автор книги «Доказательства эволюции»). Если вы не нашли необходимую информацию о книге — напишите в комментариях, мы постараемся отыскать её.
). Отличия файла от оригинала минимальны. В файле отсутствуют по понятным причинам несколько видеороликов, которые имеются в оригинале, но все изображения и гиперссылки сохранены. Кроме того, в файле отсутствует самая последняя глава — фрагменты из книги Ф. Коллинза.
Доказательства эволюции — читать онлайн бесплатно полную книгу (весь текст) целиком
Ниже представлен текст книги, разбитый по страницам. Система сохранения места последней прочитанной страницы, позволяет с удобством читать онлайн бесплатно книгу «Доказательства эволюции», без необходимости каждый раз заново искать на чём Вы остановились. Поставьте закладку, и сможете в любой момент перейти на страницу, на которой закончили чтение.
Интервал:
Закладка:
Все эти факты говорят о том, что репродуктивная изоляция, считающаяся одним из главных критериев вида (или даже самым главным), может быстро развиваться под действием отбора как в эксперименте, так и в природных условиях. Возникновение репродуктивной изоляции — ключевой этап видообразования. После того, как между генофондами двух популяций прекратился обмен генами, дальнейшее расхождение (дивергенция) этих популяций становится неизбежной просто в силу статистических факторов. В двух популяциях будут фиксироваться разные мутации, как нейтральные, так и полезные. Рано или поздно это неизбежно приведет к полной генетической несовместимости двух разошедшихся видов. Каждая новая мутация должна быть совместима с остальными генами в генофонде популяции, иначе отбор ее отсеет. Однако мутации, возникающие в одной из популяций, не проверяются отбором на совместимость с генофондом другой популяции. Следовательно, рано или поздно в популяции А закрепится мутация, несовместимая с генофондом популяции Б. В природе на формирование полной генетической несовместимости между двумя изолированными популяциями уходит, как правило, от 2–3 до 10–15 млн лет. Но уже задолго до этого момента разошедшие популяции считаются (и являются) двумя разными видами, потому что реально не скрещиваются в природе, хотя и сохраняют такую возможность.
Промежуточные этапы видообразования
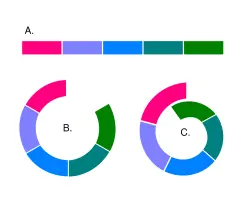
Диаграмма, демонстрирующая варианты скрещивания соседних популяций одного вида. В третьем случае распространение форм может образовать кольцо, в результате две соседние популяции друг с другом не скрещиваются, т. е. формально являются разными видами, хотя и связаны между собой непрерывным рядом взаимно совместимых популяций.
В определении понятия биологического видасущественное значение имеет практически полная репродуктивная изоляцияв природных условиях. Если видообразованиепроисходит постепенно, в результате небольших последовательных изменений, то и сегодня мы должны наблюдать все возможные стадии видообразования, а значит и все возможные стадии репродуктивной изоляции: свободное скрещивание между популяциями, затрудненное скрещивание, снижение плодовитости потомства, бесплодное потомство, и наконец — полную генетическую изоляцию. Если ничего подобного среди современных видов обнаружить не удается, то тем самым опровергаются основы эволюционного учения, то есть это пример фальсифицируемостиэволюционного учения (см.: Speciation stages).
Существует множество примеров того, как разные виды могут скрещиваться в исключительных условиях. Например, у черной вороныи серой вороныразные ареалы, но на пересечении этих ареалов виды могут образовывать гибриды. Еще один пример — различные виды рыбы рода чукучановиз реки Платт, которые живут рядом, но редко скрещиваются.
Неполное видообразование можно наблюдать у различных кольцевых видов: представители вида обитают вокруг гор, водоемов и т. п., при этом соседние популяции скрещиваются между собой и незначительно различаются, а крайние формы различаются значительно и не скрещиваются (см. диаграмму). Пример кольцевого вида — безлёгочная саламандра Ensatina eschscholtzii , обитающая на склонах гор близ тихоокеанского побережья Северной Америки. В зависимости от района обитания вокруг гор саламандры образуют различные формы, постепенно изменяющие свои морфологические и экологические характеристики. Крайние формы Ensatina eschscholtzii eschscholtzii и Ensatina eschscholtzii klauberi , первая из которых имеет красноватую окраску, а вторая чёрно-белую, сосуществуют в узкой зоне гор на юге Калифорнии, однако между собой не скрещиваются.
Еще один пример — серебристая чайка ( Larus argentatus ) и клуша ( Larus fuscus ). Они встречаются в Англии, где их признали разными видами. Но при этом между ними существует цепочка гибридных популяций, идущая вокруг северного полюса. Другие примеры кольцевых видов — зелёная пеночка ( Phylloscopus trochiloides ), оленья мышь ( Реromyscus maniculatus ), наземная улитка ( Partula olympia ), дрозофила Drosophila paulistorum .
У многих гибридов снижена фертильность. У гибридов землероек из рода бурозубок размножение затруднено вследствие разницы хромосомных наборов. Тот же эффект наблюдался в лаборатории при скрещивании Drosophila pseudoobscura из Юты и Калифорнии. Фертильность также снижена у гибридов лягушек из семейства бомбиновых жерлянки краснобрюхой (Bombina bombina) и жерлянки желтобрюхой (Bombina variegata).
Читать дальшеИнтервал:
Закладка:
Похожие книги на «Доказательства эволюции»
Представляем Вашему вниманию похожие книги на «Доказательства эволюции» списком для выбора. Мы отобрали схожую по названию и смыслу литературу в надежде предоставить читателям больше вариантов отыскать новые, интересные, ещё непрочитанные произведения.







Обсуждение, отзывы о книге «Доказательства эволюции» и просто собственные мнения читателей. Оставьте ваши комментарии, напишите, что Вы думаете о произведении, его смысле или главных героях. Укажите что конкретно понравилось, а что нет, и почему Вы так считаете.