Александр Коган - Основы физиологии высшей нервной деятельности
Здесь есть возможность читать онлайн «Александр Коган - Основы физиологии высшей нервной деятельности» весь текст электронной книги совершенно бесплатно (целиком полную версию без сокращений). В некоторых случаях можно слушать аудио, скачать через торрент в формате fb2 и присутствует краткое содержание. Город: М., Год выпуска: 1988, ISBN: 1988, Издательство: Высшая школа, Жанр: Биология, Медицина, на русском языке. Описание произведения, (предисловие) а так же отзывы посетителей доступны на портале библиотеки ЛибКат.

- Название:Основы физиологии высшей нервной деятельности
- Автор:
- Издательство:Высшая школа
- Жанр:
- Год:1988
- Город:М.
- ISBN:5-06-001444-4
- Рейтинг книги:4 / 5. Голосов: 1
-
Избранное:Добавить в избранное
- Отзывы:
-
Ваша оценка:
Основы физиологии высшей нервной деятельности: краткое содержание, описание и аннотация
Предлагаем к чтению аннотацию, описание, краткое содержание или предисловие (зависит от того, что написал сам автор книги «Основы физиологии высшей нервной деятельности»). Если вы не нашли необходимую информацию о книге — напишите в комментариях, мы постараемся отыскать её.
Максимальная ширина картинки в этой fb2-книге — 600 пикселей, но большинство картинок имеют размер не более 460 пикселей.
Основы физиологии высшей нервной деятельности — читать онлайн бесплатно полную книгу (весь текст) целиком
Ниже представлен текст книги, разбитый по страницам. Система сохранения места последней прочитанной страницы, позволяет с удобством читать онлайн бесплатно книгу «Основы физиологии высшей нервной деятельности», без необходимости каждый раз заново искать на чём Вы остановились. Поставьте закладку, и сможете в любой момент перейти на страницу, на которой закончили чтение.
Интервал:
Закладка:
Взаимодействие коры и подкорковых отделов мозга осуществляется и путем тонических влияний, регулирующих функциональное состояние нервных центров. Общеизвестно, как влияют настроение, эмоциональное состояние на эффективность умственной деятельности. И.П. Павлов говорил, что подкорка «заряжает» кору. Нейрофизиологические исследования механизмов подкорковых влияний на кору показали, что ретикулярная формация среднего мозга оказывает на нее восходящее активирующее действие. Получая коллатерали от всех афферентных путей, ретикулярная формация участвует во всех реакциях поведения, обусловливая деятельное состояние коры. Однако ее активирующее влияние при условном рефлексе организуется сигналами из проекционных зон коры (рис. 8). Раздражение ретикулярной формации вызывает изменение электроэнцефалограммы в виде ее десинхронизации, характерной для состояния активного бодрствования.
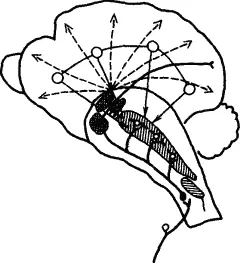
Рис. 8. Взаимодействие ретикулярной формации среднего мозга и коры (по Л.Г. Воронину):
жирными линиями обозначены афферентные специфические пути с коллатералями к ретикулярной формации, прерывистыми — восходящие пути к коре, тонкими — влияние коры на ретикулярную формацию, вертикальной штриховкой — облегчающая зона, горизонтальной — тормозная зона, клеточной штриховкой — таламические ядра
Иное действие на функциональное состояние коры оказывают специфические ядра таламуса. Их низкочастотное раздражение приводит к развитию в коре процессов торможения, которое может привести к засыпанию животного, и пр. Раздражение этих ядер вызывает в электроэнцефалограмме появление своеобразных волн — «веретен», которые переходят в медленные дельта-волны, характерные для сна. Ритм веретен может определяться тормозными постсинаптическими потенциалами (ТПСП) в нейронах гипоталамуса. Наряду с регуляторным влиянием неспецифических подкорковых структур на кору наблюдается и обратный процесс. Такие двусторонние корково-подкорковые взаимные влияния являются обязательными в осуществлении механизмов формирования временных связей.
Результаты некоторых экспериментов истолковывали как доказательство тормозящего действия стриарных структур на поведение животных. Однако дальнейшие исследования, в частности опыты с разрушением и стимуляцией хвостатых тел, и другие факты привели к заключению о наличии более сложных корково-подкорковых отношений.
Некоторые исследователи рассматривают факты об участии подкорковых структур в процессах высшей нервной деятельности как основание для того, чтобы считать их местом замыкания временных связей. Так возникло представление о «центрэнцефалической системе» как ведущей в поведении человека (У. Пенфилд, Г. Джаспер, 1958). В качестве доказательства замыкания временной связи в ретикулярной формации приводили наблюдения о том, что при выработке условного рефлекса первые изменения электрической активности мозга возникают именно в ретикулярной формации, а потом уже в коре больших полушарий. Но это свидетельствует лишь о вполне понятном раннем включении восходящей системы активации коры. Наконец, сильным аргументом в пользу подкорковой локализации замыкания считали возможность выработки условного, например зрительно-двигательного, рефлекса, несмотря на многократное рассечение коры на всю глубину, прерывающее все корковые пути между зрительной и двигательной областями. Однако этот экспериментальный факт не может служить доказательством, так как замыкание временной связи в коре имеет множественный характер и может происходить в любом ее участке между афферентными и эффекторными элементами. На рис. 9 жирными линиями показан путь условного зрительно-двигательного рефлекса при разрезах коры между зрительной и двигательной областями.
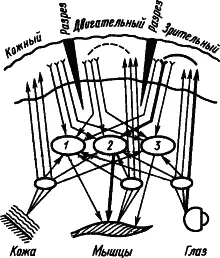
Рис. 9. Множественное замыкание временных связей в коре (показано пунктиром), которым не препятствуют ее разрезы (по А.Б. Когану):
1, 2, 3 — центральные механизмы оборонительных, пищевых и ориентировочных реакций соответственно; путь условного пищевого рефлекса на световой сигнал показан жирными линиями
Как показали многочисленные исследования, участие подкорковых структур в процессах высшей нервной деятельности не ограничивается регуляторной ролью ретикулярной формации среднего мозга и лимбических структур. Ведь уже на подкорковом уровне происходят анализ и синтез действующих раздражителей и оценка их биологического значения, в значительной мере определяющая характер образующихся связей с сигналом. Использование показателей формирования наиболее коротких путей, по которым сигнал достигает разных подкорковых структур мозга, выявило наиболее выраженное участие в процессах обучения задних отделов таламуса и поля CA 3гиппокампа. Роль гиппокампа в явлениях памяти подтверждается многими фактами. Наконец, нет оснований предполагать, что способность к примитивной замыкательной деятельности структур мозга, которая приобреталась в эволюции, когда они были ведущими, теперь у них полностью исчезла, когда эта функция перешла к новой коре.
Читать дальшеИнтервал:
Закладка:
Похожие книги на «Основы физиологии высшей нервной деятельности»
Представляем Вашему вниманию похожие книги на «Основы физиологии высшей нервной деятельности» списком для выбора. Мы отобрали схожую по названию и смыслу литературу в надежде предоставить читателям больше вариантов отыскать новые, интересные, ещё непрочитанные произведения.










Обсуждение, отзывы о книге «Основы физиологии высшей нервной деятельности» и просто собственные мнения читателей. Оставьте ваши комментарии, напишите, что Вы думаете о произведении, его смысле или главных героях. Укажите что конкретно понравилось, а что нет, и почему Вы так считаете.