Владимир Комаров - Происхождение растений
Здесь есть возможность читать онлайн «Владимир Комаров - Происхождение растений» — ознакомительный отрывок электронной книги совершенно бесплатно, а после прочтения отрывка купить полную версию. В некоторых случаях можно слушать аудио, скачать через торрент в формате fb2 и присутствует краткое содержание. Город: Москва, Год выпуска: 1961, Издательство: Издательство Академии наук СССР, Жанр: Биология, sci_botany, на русском языке. Описание произведения, (предисловие) а так же отзывы посетителей доступны на портале библиотеки ЛибКат.

- Название:Происхождение растений
- Автор:
- Издательство:Издательство Академии наук СССР
- Жанр:
- Год:1961
- Город:Москва
- ISBN:нет данных
- Рейтинг книги:3 / 5. Голосов: 1
-
Избранное:Добавить в избранное
- Отзывы:
-
Ваша оценка:
Происхождение растений: краткое содержание, описание и аннотация
Предлагаем к чтению аннотацию, описание, краткое содержание или предисловие (зависит от того, что написал сам автор книги «Происхождение растений»). Если вы не нашли необходимую информацию о книге — напишите в комментариях, мы постараемся отыскать её.
Происхождение растений — читать онлайн ознакомительный отрывок
Ниже представлен текст книги, разбитый по страницам. Система сохранения места последней прочитанной страницы, позволяет с удобством читать онлайн бесплатно книгу «Происхождение растений», без необходимости каждый раз заново искать на чём Вы остановились. Поставьте закладку, и сможете в любой момент перейти на страницу, на которой закончили чтение.
Интервал:
Закладка:
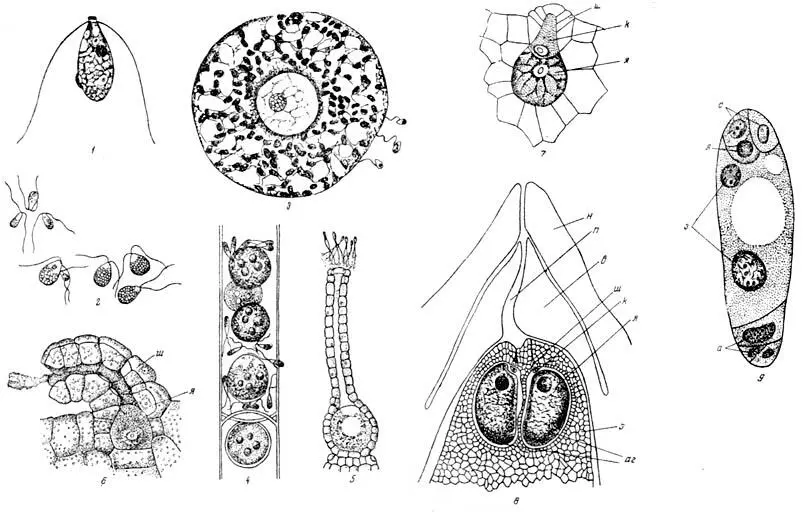
Рис. 26. Развитие гаплоидных элементов от изогамет до зародышевого ядра цветковых растений
1 — изогамета водоросли «морокой салат» (Ulva latuca); 2 — гетерогамия: макро- и микрогаметы водоросли Codium; 3 — яйцеклетка водоросли диктиота в момент оплодотворения; справа — микрогаметы (сперматозоиды); 4 — оогоний водоросли (сперматозоиды); 5 — архегоний мха с яйцеклеткой в нижней, расширенной части сфероплеа с несколькими яйцеклетками и проникшими в него извне микрогаметами и сперматозоидами у входа в его шейку наверху; 6 — архегоний папоротника, несколько упрощенный по сравнению с архегонием мха; яйцеклетка частично погружена в ткань заростка. Из канала шейки выходит слизь, получившаяся при расплывании канальцевых клеток; 7 — еще более упрощенный архегоний папоротника сальвиния; 8 — архегоний сосны, находящиеся среди ткани заростка в зародышевом мешке, внутри семяпочки; 9 — зародышевый мешок лилии, вынутый из семяпочки; н — наружный покров семяпочки; п — канал пыльцевой камеры; в — внутренний покров семяпочки; э — ткань заростка, превращенная в эндосперм; аг — два архегония; ш — шейка архегония; к — канальцевая клетка; о — антиподы (ткань заростка); з — ядра зародышевого мешка, из которых после оплодотворения возникает эндосперм; с — спутник или синэргиды; я — яйцеклетка
Мы не знаем, как развивались псилофиты, по у современных мхов мы находим в пазухах листьев, часто собранными помногу на концах веточек, еще более сложные органы архегонии. Архегоний состоит из шейки и брюшка, стенка его многоклетная, полость брюшка заполнена крупной мощной яйцеклеткой или иначе макрогаметой; канал шейки выполнен несколькими легко расплывающимися клетками. Проникающий сквозь канал сперматозоид вызывает оплодотворение, связанное, как и всегда, с удвоением числа хромосом. Яйцеклетка превращается в зиготу, последняя делится и образует своим дальнейшим ростом палочкообразный зародыш, из которого позднее образуется спорогон мха.
У мхов, хвощей, настоящих папоротников и некоторых других высших споровых все споры одинаковы, и угадать, вырастет ли из нее гаметофит, способный дать и макро- и микрогаметангии, или же только макрогаметангии — архегонии, или же только микрогаметангии — антеридии, невозможно.
Между тем уже у крупных деревьев девонского и каменноугольного периодов, именно у лепидодендронов, было полное разделение на макро- и микроспоры. Из макроспор вырастали макрогаметофиты, т. е. заростки, дававшие архегонии и в них яйцеклетки. Английскому палеонтологу Макину (Maclean) удалось получить тонкие шлифы через макроспоры, на которых хорошо видно, как образовавшийся уже в макроспоре заросток своей верхней частью выдается из лопнувшей макроспоры, образуя тонкую, вероятно, зеленую волнистую пластинку, на которой сидят архегонии. Большая часть этого заростка, вероятно бесцветная, оставалась внутри споры и состояла из ткани, богатой запасными питательными веществами.
У современных нам плауновых, называемых селагинеллами, у маленькой группы шильников (Isoetaceae), у некоторых водяных папоротников (Salviniaceae и Marsiliaceae) деление на макро- и микроспоры выражено очень хорошо и позволяет во всех деталях проследить развитие мужских и женских заростков. Яйцеклетка у них находится в архегонии, который погружен в ткань заростка. Если бы заросток не выступал из макроспоры, а развивался бы целиком внутри ее, то защита первых стадий зародышевой жизни растения была бы еще более обеспечена, что мы и видим у цветковых растений.
У голосеменных растений редукция (упрощение) гаметофита идет значительно далее. Архегонии состоят из крупной яйцеклетки, на верхушке которой сидит несколько маленьких клеточек, сохранившихся как рудимент шейки архегония.
У голосеменных и цветковых растений, как возникших в процессе борьбы за овладение засушливыми территориями юрских материков, гаметофит и яйцевая клетка глубоко запрятаны под защитой тканей спорофита. Макроспоры возникают путем редукционного деления в глубине тканей семяпочки, в полости завязи, окруженной покровами цветка или соцветия. Из четырех макроспор зрелости достигает только одна, она сильно разрастается и превращается в так называемый зародышевый мешок, крупную клетку с двумя гаплоидными ядрами в центре. Прорастание этой макроспоры и развитие в ней заростка состоят в том, что у верхнего ее конца развиваются три крупных ядра — зародышевое и два его спутника (две синэргиды), а у нижнего три клетки, называемые антиподами. Последние можно рассматривать как вегетативную ткань заростка. Считая и центральные ядра, всего в зародышевом мешке развивается 8 ядер. В отдельных случаях число это значительно больше, так как антипод может быть много.
Читать дальшеИнтервал:
Закладка:
Похожие книги на «Происхождение растений»
Представляем Вашему вниманию похожие книги на «Происхождение растений» списком для выбора. Мы отобрали схожую по названию и смыслу литературу в надежде предоставить читателям больше вариантов отыскать новые, интересные, ещё непрочитанные произведения.












Обсуждение, отзывы о книге «Происхождение растений» и просто собственные мнения читателей. Оставьте ваши комментарии, напишите, что Вы думаете о произведении, его смысле или главных героях. Укажите что конкретно понравилось, а что нет, и почему Вы так считаете.