БСЭ БСЭ - Большая Советская Энциклопедия (РИ)
Здесь есть возможность читать онлайн «БСЭ БСЭ - Большая Советская Энциклопедия (РИ)» весь текст электронной книги совершенно бесплатно (целиком полную версию без сокращений). В некоторых случаях можно слушать аудио, скачать через торрент в формате fb2 и присутствует краткое содержание. Жанр: Энциклопедии, на русском языке. Описание произведения, (предисловие) а так же отзывы посетителей доступны на портале библиотеки ЛибКат.

- Название:Большая Советская Энциклопедия (РИ)
- Автор:
- Жанр:
- Год:неизвестен
- ISBN:нет данных
- Рейтинг книги:4 / 5. Голосов: 1
-
Избранное:Добавить в избранное
- Отзывы:
-
Ваша оценка:
Большая Советская Энциклопедия (РИ): краткое содержание, описание и аннотация
Предлагаем к чтению аннотацию, описание, краткое содержание или предисловие (зависит от того, что написал сам автор книги «Большая Советская Энциклопедия (РИ)»). Если вы не нашли необходимую информацию о книге — напишите в комментариях, мы постараемся отыскать её.
Большая Советская Энциклопедия (РИ) — читать онлайн бесплатно полную книгу (весь текст) целиком
Ниже представлен текст книги, разбитый по страницам. Система сохранения места последней прочитанной страницы, позволяет с удобством читать онлайн бесплатно книгу «Большая Советская Энциклопедия (РИ)», без необходимости каждый раз заново искать на чём Вы остановились. Поставьте закладку, и сможете в любой момент перейти на страницу, на которой закончили чтение.
Интервал:
Закладка:
Транспортные Р. к. (тРНК): имеют константу седиментации около 4 S и молекулярную массу около 25 000, т. е. являются сравнительно низкополимерными (около 80 нуклеотидных остатков); содержат относительно много метилированных и других минорных оснований. Их биологическая роль заключается в присоединении активированных аминокислотных остатков и переносе (транспорте) их на рибосомы, т. е. к месту синтеза полипептидных цепочек. Для каждой аминокислоты имеются свои специфические тРНК (обычно более одной). Все тРНК имеют сложную, частью двуспиральную, макромолекулярную структуру, изображаемую в виде клеверного листа. Они содержат участки, присоединяющиеся к рибосоме, триплет нуклеотидов ( антикодон ) , присоединяющийся к кодону мРНК, и концевой участок, присоединяющий аминокислотный остаток. Первичная структура более 60 тРНК установлена полностью.
Информационные, или матричные, Р. к. (иРНК, или мРНК) представляют собой наиболее разнородную группу и играют роль матриц при биосинтезе белков в процессе трансляции (считывания нуклеотидного кода и перевода его в определённую последовательность аминокислот в полипептидных цепях белков). Все виды Р. к. синтезируются в клетках на матрице ДНК, образуя последовательность рибонуклеотидов, комплементарную последовательности дезоксирибонуклеотидов в ДНК (процесс транскрипции). В клеточном ядре обнаружены гигантские молекулы — предшественники мРНК, большая часть которых распадается внутри ядра и только сравнительно небольшая часть молекулы переходит в цитоплазму и образует собственно мРНК. Быстро распадающаяся в клеточном ядре Р. к., вероятно, играет регуляторную роль. Обнаружены также некоторые др. типы Р. к., например низкополимерная относительно стабильная ядерная РНК, роль которой пока неясна. См. также статьи Белки, Молекулярная генетика, Нуклеиновые кислоты, Рибосомы и лит. при них.
Лит.: Дэвидсон Дж., Биохимия нуклеиновых кислот, пер. с англ., М., 1968; Химия и биохимия нуклеиновых кислот, под ред. И. Б. Збарского и С. С. Дебова, Л., 1968; Спирин А. С. и Гаврилов а Л. П., Рибосома, 2 изд., М., 1971; Darnell J., Ribonucleis acids from animal cells, «Bacteriological Reviews», 1968, v. 32, № 3, p. 262; Attardi G. and Amaid i F., Structure and synthesis of ribosomal RNA, «Annual Review of Biochemistry», 1970, v. 39, p. 183; Wienberg R. A., Nuclear RNA metabolism, «Annual Review of Biochemistry», 1973, v. 42, p. 329.
И. Б. Збарский.
Рибосомы
Рибосо'мы,внутриклеточные частицы, осуществляющие биосинтез белка; Р. обнаружены в клетках всех без исключения живых организмов: бактерий, растений и животных; каждая клетка содержит тысячи или десятки тысяч Р.
Форма Р. близка к сферической, хотя её очертания сложны и не могут быть описаны простой геометрической фигурой. Различают 2 главных класса Р.: так называемые 70 SP (молекулярная масса около 3×10 6, диаметром около 200—300 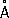 , коэффициент седиментации S° 20wоколо 70 единиц Сведберга) и более крупные 80 S P. (молекулярная масса около 4—5×10 6, максимальный размер до 400
, коэффициент седиментации S° 20wоколо 70 единиц Сведберга) и более крупные 80 S P. (молекулярная масса около 4—5×10 6, максимальный размер до 400  , коэффициент седиментации около 80 единиц Сведберга). Р. 70 S класса характерны для клеток, не имеющих оформленного ядра, — прокариотов (бактерии, актиномицеты и синезелёные водоросли), а также для хлоропластов и митохондрий высших организмов. Р. 80 S класса обнаружены в цитоплазме всех эукариотов, т. е. организмов, имеющих оформленное клеточное ядро. По химической природе Р. — нуклеопротеид, состоящий из рибонуклеиновой кислоты (РНК) и белка. Р. класса 70 S содержит 60—65% РНК и 40—35% белка, а Р. класса 80 S — около 50% РНК и 50% белка. Универсальный принцип структурной организации Р. — построение её из двух неравных субчастиц (субъединиц), на которые она может диссоциировать (например, при понижении концентрации ионов Mg 2+в среде) и вновь реассоциировать по схеме:
, коэффициент седиментации около 80 единиц Сведберга). Р. 70 S класса характерны для клеток, не имеющих оформленного ядра, — прокариотов (бактерии, актиномицеты и синезелёные водоросли), а также для хлоропластов и митохондрий высших организмов. Р. 80 S класса обнаружены в цитоплазме всех эукариотов, т. е. организмов, имеющих оформленное клеточное ядро. По химической природе Р. — нуклеопротеид, состоящий из рибонуклеиновой кислоты (РНК) и белка. Р. класса 70 S содержит 60—65% РНК и 40—35% белка, а Р. класса 80 S — около 50% РНК и 50% белка. Универсальный принцип структурной организации Р. — построение её из двух неравных субчастиц (субъединиц), на которые она может диссоциировать (например, при понижении концентрации ионов Mg 2+в среде) и вновь реассоциировать по схеме:
70 S Û 50 S + 30 S; 80 S Û 60 S + 40 S
Большая субчастица (50 S или 60 S) состоит из молекулы высокополимерной рибосомальной РНК (молекулярная масса 1,1—1,8×10 6), молекулы относительно низкополимерной рибосомальной РНК (молекулярная масса 40 000) и нескольких десятков молекул белков. Малая субчастица (30 S или 40 S) состоит из молекулы высокополимерной рибосомальной РНК (молекулярная масса 0,6—0,7×10 6) и от 20 (в 30 S частицах) до 40 (в 40 S частицах) различных молекул белков. Высокополимерная рибосомальная РНК создаёт возможность сборки этих белков в единую рибонуклеопротеидную частицу. В эксперименте можно осуществить разворачивание Р.: частица становится более рыхлой, РНК разворачивается в тяж, при этом все белки остаются связанными с ней. В других условиях можно добиться последовательного отделения белков от РНК (разборка Р.). Эта разборка обратима, и в подходящих условиях белки и РНК снова спонтанно объединяются в рибонуклеопротеид, формирующий нативную структуру Р. (самосборка Р.). Считают, что образование Р. в клетках также идёт путём самосборки из предварительно синтезированных РНК и белков.
Читать дальшеИнтервал:
Закладка:
Обсуждение, отзывы о книге «Большая Советская Энциклопедия (РИ)» и просто собственные мнения читателей. Оставьте ваши комментарии, напишите, что Вы думаете о произведении, его смысле или главных героях. Укажите что конкретно понравилось, а что нет, и почему Вы так считаете.