Richard Powers - Gold Bug Variations
Здесь есть возможность читать онлайн «Richard Powers - Gold Bug Variations» весь текст электронной книги совершенно бесплатно (целиком полную версию без сокращений). В некоторых случаях можно слушать аудио, скачать через торрент в формате fb2 и присутствует краткое содержание. Год выпуска: 1991, Издательство: Harper Perennial, Жанр: Современная проза, на английском языке. Описание произведения, (предисловие) а так же отзывы посетителей доступны на портале библиотеки ЛибКат.

- Название:Gold Bug Variations
- Автор:
- Издательство:Harper Perennial
- Жанр:
- Год:1991
- ISBN:нет данных
- Рейтинг книги:5 / 5. Голосов: 1
-
Избранное:Добавить в избранное
- Отзывы:
-
Ваша оценка:
Gold Bug Variations: краткое содержание, описание и аннотация
Предлагаем к чтению аннотацию, описание, краткое содержание или предисловие (зависит от того, что написал сам автор книги «Gold Bug Variations»). Если вы не нашли необходимую информацию о книге — напишите в комментариях, мы постараемся отыскать её.
Gold Bug Variations — читать онлайн бесплатно полную книгу (весь текст) целиком
Ниже представлен текст книги, разбитый по страницам. Система сохранения места последней прочитанной страницы, позволяет с удобством читать онлайн бесплатно книгу «Gold Bug Variations», без необходимости каждый раз заново искать на чём Вы остановились. Поставьте закладку, и сможете в любой момент перейти на страницу, на которой закончили чтение.
Интервал:
Закладка:
He slips out of the party, the mocked-up festive lab. He stands in the darkened hall, a hundred steps down, in a blind recess, waiting. Five minutes turns into an agonized ten. Surely she must have seen him leave. At last, she hurries out furtively, looking over her shoulder in fear, sheer erotic terror at being caught. He steps from his shadow. She stifles a shout and collapses into him, clinging.
"Listen," he orders. "Nature, 1955. Gale and Folkes. Test-tube protein synthesis. Incorporation. I told Tooney, before he left. He thinks it'll work. We place the sequence to study in glass. Out comes the offending enzyme."
"Shh," she says, convulsing rhythmically. "I love you." The sound of singing, candle scent from the far end of the hall. He holds her to him, all along his length. Her tangled hair, her face, her muscular shoulders, the small of her back, her upper legs. "Make up for lost time," she laughs, sniffling. She lets out a short, soft, pained cry midway between a howler monkey and a gothic angel's et exultavit. He signals her, unnecessarily in the dark: Don't even say it.
Deus ex Machina
Q: Who made me? Defensible evidence only please.
A molecule able to influence two others that would not react otherwise: can my miracle reside here? Does DNA, the map unfolding the whole organism, do no more than manufacture reagents, golem formulae, tinctures where soul emerges if the secret proportions are hit on? The code I am after must embody not just stuff but substance: process, decision, feedback. Not data alone; behavior at molecular level.
The lint-ball tangle of an enzyme — its charged terrain of twists and turns, vise-grips for welding chemical substrate — makes it a three-dimensional, supple machine. Here is the muse of fire I've been needing. Certain of these enzyme proteins become single-molecule transistors, devices that test and respond to feedback, creating a free repertoire from predictable physics. The assembled amino string of an allosteric enzyme can tangle into two different shapes. With unique twists in each shape, it thus possesses two separate sets of binding sites. The molecule may be enzymatically active in one shape and inert in the other, like a shoehorn that sometimes warps into worthlessness. A substance that binds to a site in either the active or inert shape will lock the enzyme into that configuration:
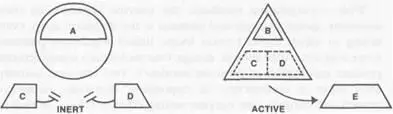
The inert shape of this enzyme has a binding site that fits substance A. The active shape has a site matching B as well as materials C and D that it transforms into product E. If A grabs the molecule first, it locks it into inert shape, eliminating those sites that accept the catalyzable materials. The enzyme is switched off, C and D can't bind, and manufacture of E stops. But if B first binds to the enzyme in active form, it locks the molecule into a shape with C and D's sites intact. The faucet is held open; the enzyme joins C and D into E so long as supplies of C and D exist.
Ressler's magic Boolean circuitry begins to emerge. The presence of A inhibits the manufacture of E; B promotes it. None of the compounds reacts with the enzyme itself; the machine remains unchanged except for switching on or off, always capable of switching back if the splint-substances detach. Even wilder: the inhibitors, promoters, and inputs, binding to independent sites, need have nothing to do with one another. A, B, C, D, and E can be anything at all. In theory, any chemical can be made to inhibit or promote the formation or degradation of any other. The effect can even be nonlinear; multiple binding sites on an enzyme could cause small amounts of compound to have enormous effects on the synthesis of others.
Here it is, my escape act from chemical necessity. Microcircuitry I can't begin to map: a single allosteric enzyme made up of a few hundred amino acids, weighing less than a million billionth of a gram, accepting multiple, graded inputs and producing nonlinear output, a free-floating if-then program. It smacks of religion to me.
But the conjuring act hasn't even begun. Link the logical feeds of an allosteric circuit together, and the molecule virtually lives. E, the reaction product, can be the same as A, the inhibitor. Every successful catalysis then shuts off the switch. Or E, used up by the body, might degrade into promoter B, ensuring that new E is created whenever old stock is expended.
With self-regulating feedback, the enzyme becomes its own economy, gauging supply and demand in the chemical soup, even acting to adjust these. I enter loops, linked regulatory patterns more ingenious than theistic design. One molecule's manufactured product can inhibit or promote another's. Two enzymes activate each other in conjunction or opposition. Metabolic pathways branch and conjoin, one enzyme setting off two others, or two in tandem combining to shut down a first. These and, or, and not operations create a complete prepositional calculus.
Coordinated microprograms capable of changing their own environment, able to spring into production modes within an instant of encountering a trigger, create a cybernetic network powerful enough to initiate the impossibly articulated behavior of the composite cell. Q: Is the tracery of microprogramming networks too complex to have arisen through guided chance? Q: Is it complex enough to account for the autonomy that high-level enzyme byproducts — Ressler, Todd, myself — all suffer from?
I've started four times today, on four separate sheets — what? Nothing. Trivial message strings going no farther than Dear Franklin. Even the adjective is problematic. After months, I have the man's address. I have a world of things to tell him. Nothing stops me. I want to write. But even today — Caesar crosses Rubicon, 49 B.C. — I can't. I won't. English doesn't have the modality to say what's keeping me. Writing him is fine, but words are out. A is too functional, B too forgiving. C and D are transparent excuses for E, which I will never bring myself to say to him again.
I own his mailbox, the lookup table to the one spot where he can be reached. If I could finish a fifth sheet, seal and mail it without reading… I could; I want to. The first letter of the first sentence, and I waver from one urge to the other. I am truly stochastic, indecisive. Do all inputs, computed, already drive me to one course or the other, or can they still be interfered with by some messy conglomerate circuit, me?
I try on Laplace's old dream — to solve the world through giant inference engine. Ressler, alert, talkative as I had never before seen him, racing through the data stacks looking for our place to cut, cracked a joke about the final triumph of the reductionist program. "NASA's eyes in the sky determine the vectors on every molecule of atmosphere. They feed all these numbers into a Cray, and the animal pounds away, megaflops, on a simulation that knows everything about adiabatic cooling, turbulence, vapor pressures, topography, solar radiation. The machine assembles and delivers a perfect prediction of tomorrow's weather. Only it takes two days to run."
The problem is irresistible. Do all my active enzymes plus the running average of the chemical soup they find themselves in, the jungle of bioeconomy (vast, uncatalogued tracts of electrochemical memory, mine and earlier), all the stimuli bombarding me from outside — the January sun's false springs out my window, the glass of water at my elbow (complete with Brooklyn reservoir heavy impurities), the feel of the keys under my fingers — do all these independent effectors sum to one unique output: write him or not? They couldn't sum to more than one outcome. What would it even mean to say the choice, the final cybernetic weighing were left open? Open to what? Whom?
Читать дальшеИнтервал:
Закладка:
Похожие книги на «Gold Bug Variations»
Представляем Вашему вниманию похожие книги на «Gold Bug Variations» списком для выбора. Мы отобрали схожую по названию и смыслу литературу в надежде предоставить читателям больше вариантов отыскать новые, интересные, ещё непрочитанные произведения.












Обсуждение, отзывы о книге «Gold Bug Variations» и просто собственные мнения читателей. Оставьте ваши комментарии, напишите, что Вы думаете о произведении, его смысле или главных героях. Укажите что конкретно понравилось, а что нет, и почему Вы так считаете.