Jane Flint - Principles of Virology, Volume 1
Здесь есть возможность читать онлайн «Jane Flint - Principles of Virology, Volume 1» — ознакомительный отрывок электронной книги совершенно бесплатно, а после прочтения отрывка купить полную версию. В некоторых случаях можно слушать аудио, скачать через торрент в формате fb2 и присутствует краткое содержание. Жанр: unrecognised, на английском языке. Описание произведения, (предисловие) а так же отзывы посетителей доступны на портале библиотеки ЛибКат.

- Название:Principles of Virology, Volume 1
- Автор:
- Жанр:
- Год:неизвестен
- ISBN:нет данных
- Рейтинг книги:5 / 5. Голосов: 1
-
Избранное:Добавить в избранное
- Отзывы:
-
Ваша оценка:
Principles of Virology, Volume 1: краткое содержание, описание и аннотация
Предлагаем к чтению аннотацию, описание, краткое содержание или предисловие (зависит от того, что написал сам автор книги «Principles of Virology, Volume 1»). Если вы не нашли необходимую информацию о книге — напишите в комментариях, мы постараемся отыскать её.
Volume I: Molecular Biology
Volume II: Pathogenesis and Control
Principles of Virology, Fifth Edition
Principles of Virology, Volume 1 — читать онлайн ознакомительный отрывок
Ниже представлен текст книги, разбитый по страницам. Система сохранения места последней прочитанной страницы, позволяет с удобством читать онлайн бесплатно книгу «Principles of Virology, Volume 1», без необходимости каждый раз заново искать на чём Вы остановились. Поставьте закладку, и сможете в любой момент перейти на страницу, на которой закончили чтение.
Интервал:
Закладка:
BOX 3.2
TERMINOLOGY
Important conventions: plus (+) and minus (–) strands
mRNA is defined as the positive (+) strand, because it can be translated. A strand of DNA of the equivalent polarity is also designated as a (+) strand; i.e., if it were mRNA, it would be translated into protein.
The RNA or DNA complement of the (+) strand is called the (–) strand. The (–) strand cannot be translated; it must first be copied to make the (+) strand. Ambisense RNA contains both (+) and (–) sequences.
A color key for nucleic acids, proteins, membranes, cells, and more is located in the front of this book.

DNA Genomes
The strategy of having DNA as a viral genome appears at first glance to be the ultimate in genetic efficiency: the host genetic system is based on DNA, so viral genome replication and expression could simply emulate the host system. While the replication of viral and cellular DNA genomes is fundamentally similar, the mechanistic details are varied because viral genomes are structurally diverse.
Double-Stranded DNA (dsDNA) (Fig. 3.2)
There are 38 families of viruses with dsDNA genomes. Those that include vertebrate viruses are the Adenoviridae , Alloherpesviridae , Asfarviridae , Herpesviridae , Papillomaviridae , Polyomaviridae , Iridoviridae , and Poxviridae . These genomes may be linear or circular. Genome replication and mRNA synthesis are accomplished by host or viral DNA-dependent DNA and RNA polymerases.
Gapped DNA (Fig. 3.3)
Members of two virus families, Caulimoviridae and He- padnaviridae , have a gapped DNA genome. The Hepadnaviridae include viruses that infect vertebrates. As the gapped DNA genome is partially double stranded, the gaps must be filled to produce perfect duplexes. This repair process must precede mRNA synthesis because the host RNA polymerase can transcribe only fully dsDNA. The unusual gapped DNA genome is produced from an RNA template by a virus-encoded enzyme, reverse transcriptase.
Single-Stranded DNA (ssDNA) (Fig. 3.4)
Thirteen families of viruses containing ssDNA genomes have been recognized; the families Anelloviridae , Circoviridae , Genomoviridae , and Parvoviridae include viruses that infect vertebrates. ssDNA must be copied into mRNA before proteins can be produced. However, RNA can be made only from a dsDNA template, whatever the sense of the ssDNA. Consequently, DNA synthesis mustprecede mRNA production in the replication cycles of these viruses. All synthesis of viral DNA is catalyzed by cellular DNA polymerases.
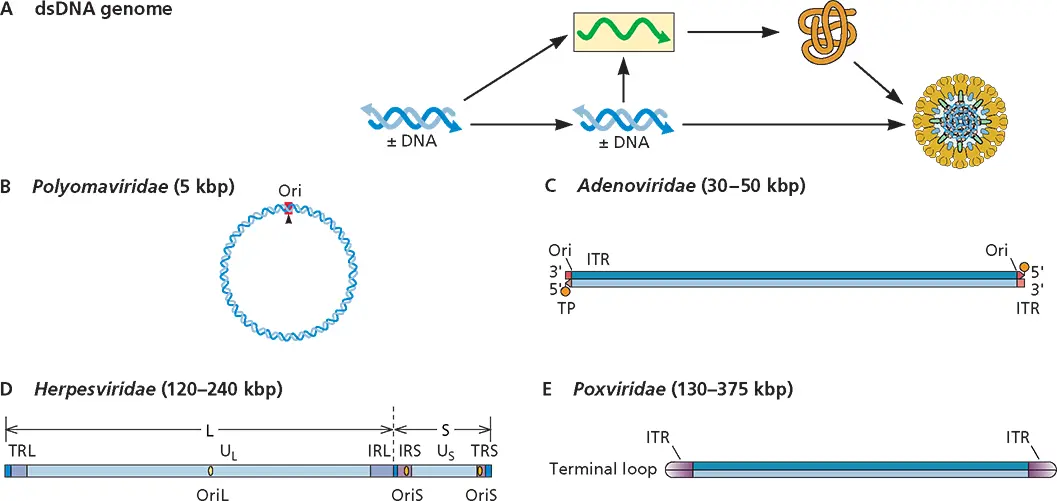
Figure 3.2 Structure and expression of viral double-stranded DNA genomes. (A)Synthesis of genomes, mRNA (shown as green line in yellow box), and protein (shown as brown line). The icon represents a polyomavirus particle. (B to E)Genome configurations. Ori, origin of replication; ITR, inverted terminal repeat; TP, terminal protein; L, long region; S, short region; U Land U S, long and short unique regions; IRL, internal repeat sequence, long region; IRS, internal repeat sequence, short region; TRL, terminal repeat sequence, long region; TRS, terminal repeat sequence, short region; OriL, origin of replication of the long region; OriS, origin of replication of the short region.
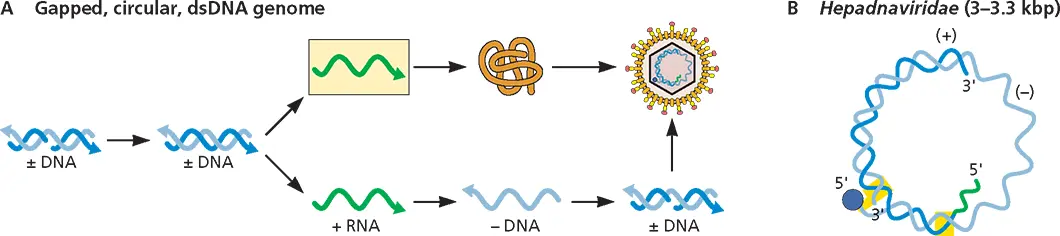
Figure 3.3Structure and expression of viral gapped, circular, double-stranded DNA genomes. (A)Synthesis of genome, mRNA, and protein. (B)Configuration of the hepadnavirus genome.
RNA Genomes
Cells have no RNA-dependent RNA polymerases that can replicate the genomes of RNA viruses or make mRNA from RNA templates ( Box 3.3). One solution to this problem is that RNA virus genomes encode RNA-dependent RNA polymerases that produce RNA from RNA templates. The other solution, exemplified by retrovirus genomes, is reverse transcription of the genome to dsDNA, which can be transcribed by host RNA polymerase.
dsRNA (Fig. 3.5)
There are twelve families of viruses with linear dsRNA genomes. The number of dsRNA segments in the virus particle may be 1 ( Totiviridae, Hypoviridae , and Endornaviridae , viruses of fungi, protozoa, and plants); 2 ( Partitiviridae , Birnaviridae , and Megabirnaviridae , viruses of fungi, plants, insects, fish, and chickens); 3 ( Cystoviridae , viruses of Pseudomonas bacteria); 4 ( Chrysoviridae , viruses of fungi); or 10 to 12 ( Reoviridae , viruses of protozoa, fungi, invertebrates, plants, and vertebrates). While dsRNA contains a (+) strand, it cannot be translated to synthesize viral proteins as part of a duplex. The (–) strand of the genomic dsRNA is first copied into mRNAs by a viral RNA-dependent RNA polymerase. Newly synthesized mRNAs are encapsidated and then copied to produce dsRNAs.
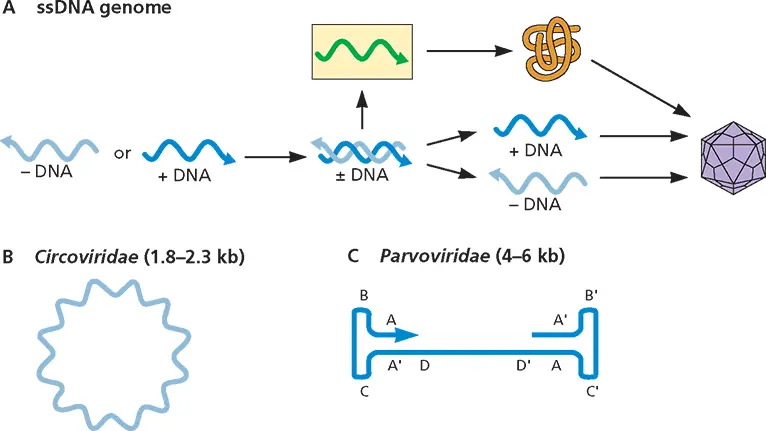
Figure 3.4Structure and expression of viral single-stranded DNA genomes. (A)Synthesis of genomes, mRNA, and protein. (B and C)Genome configurations.
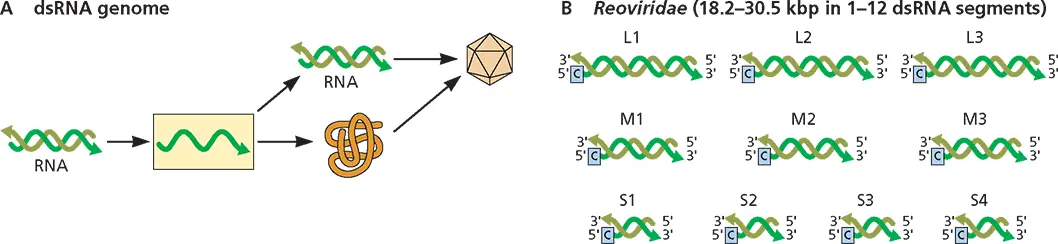
Figure 3.5Structure and expression of viral double-stranded RNA genomes. (A)Synthesis of genomes, mRNA, and protein. (B)Genome configuration.
BOX 3.3
BACKGROUND
RNA synthesis in cells
There are no known host cell enzymes that can copy the genomes of RNA viruses. However, at least one enzyme, RNA polymerase II, can copy an RNA template. The 1.7-kb circular, ssRNA genome of hepatitis delta satellite virus is copied by RNA polymerase II to form multimeric RNAs (see the figure). How RNA polymerase II, an enzyme that produces pre-mRNAs from DNA templates, is reprogrammed to copy a circular RNA template is not known.
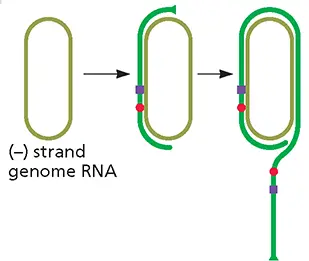
Hepatitis delta satellite (– ) strand genome RNA is copied by RNA polymerase II at the indicated position.The polymerase passes the poly(A) signal (purple box) and the self-cleavage domain (red circle). For more information, see Fig. 6.25 Redrawn from Taylor JM. 1999. Curr Top Microbiol Immunol 239:107–122, with permission.
Интервал:
Закладка:
Похожие книги на «Principles of Virology, Volume 1»
Представляем Вашему вниманию похожие книги на «Principles of Virology, Volume 1» списком для выбора. Мы отобрали схожую по названию и смыслу литературу в надежде предоставить читателям больше вариантов отыскать новые, интересные, ещё непрочитанные произведения.



Обсуждение, отзывы о книге «Principles of Virology, Volume 1» и просто собственные мнения читателей. Оставьте ваши комментарии, напишите, что Вы думаете о произведении, его смысле или главных героях. Укажите что конкретно понравилось, а что нет, и почему Вы так считаете.