Martinez J. Hewlett - Basic Virology
Здесь есть возможность читать онлайн «Martinez J. Hewlett - Basic Virology» — ознакомительный отрывок электронной книги совершенно бесплатно, а после прочтения отрывка купить полную версию. В некоторых случаях можно слушать аудио, скачать через торрент в формате fb2 и присутствует краткое содержание. Жанр: unrecognised, на английском языке. Описание произведения, (предисловие) а так же отзывы посетителей доступны на портале библиотеки ЛибКат.

- Название:Basic Virology
- Автор:
- Жанр:
- Год:неизвестен
- ISBN:нет данных
- Рейтинг книги:3 / 5. Голосов: 1
-
Избранное:Добавить в избранное
- Отзывы:
-
Ваша оценка:
Basic Virology: краткое содержание, описание и аннотация
Предлагаем к чтению аннотацию, описание, краткое содержание или предисловие (зависит от того, что написал сам автор книги «Basic Virology»). Если вы не нашли необходимую информацию о книге — напишите в комментариях, мы постараемся отыскать её.
The basics of virology Virological techniques Molecular biology Pathogenesis of human viral disease The 4th edition includes new information on the SARS, MERS and COVID-19 coronaviruses, hepatitis C virus, influenza virus, as well as HIV and Ebola. New virological techniques including bioinformatics and advances in viral therapies for human disease are also explored in-depth. The book also includes entirely new sections on metapneumoviruses, dengue virus, and the chikungunya virus.
Basic Virology — читать онлайн ознакомительный отрывок
Ниже представлен текст книги, разбитый по страницам. Система сохранения места последней прочитанной страницы, позволяет с удобством читать онлайн бесплатно книгу «Basic Virology», без необходимости каждый раз заново искать на чём Вы остановились. Поставьте закладку, и сможете в любой момент перейти на страницу, на которой закончили чтение.
Интервал:
Закладка:
The dimensions and organization of “typical” animal, plant, and bacterial cells are shown in Figure 2.1. The size of a typical virus falls in the range between the diameters of a ribosome and of a centriolar filament. With most viruses, infection of a cell with a single virus particle will result in the synthesis of more than one (often by a factor of several powers of 10) infectious virus. Any infection that results in the production of more infectious virus at the end than at the start is classified as a productive infection. The actual number of infectious viruses produced in an infected cell is called the burst size, and this number can range from less than 10 to over 10 000, depending on the type of cell infected, the nature of the virus, and many other factors.
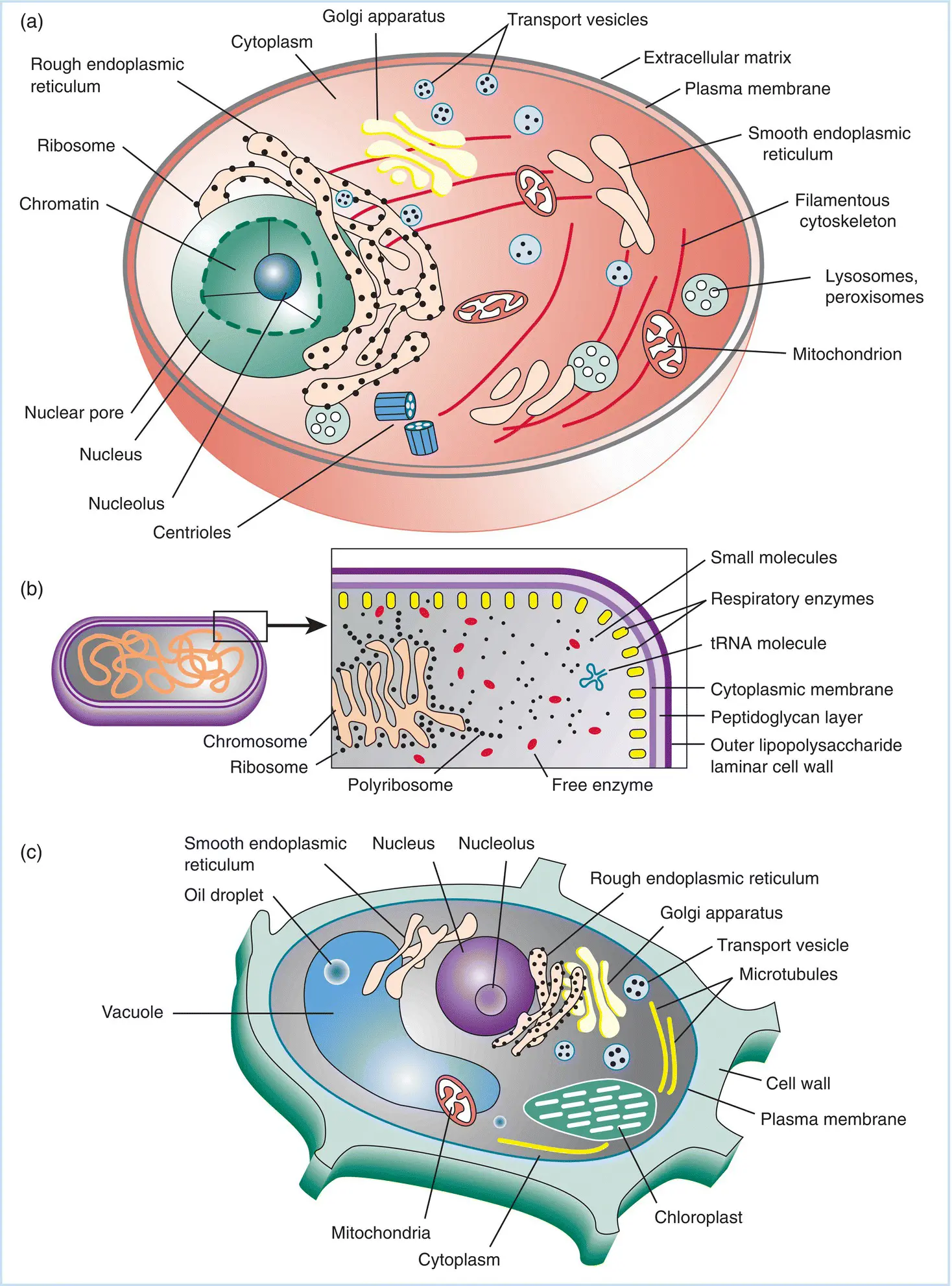
Figure 2.1Dimensions and features of “typical” (a) animal, (b) plant, and (c) bacterial cells. The dimensions of plant and animal cells can vary widely, but an average diameter of around 50 μm (5 × 10 −5m) is a fair estimate. Bacterial cells also show great variation in size and shape, but the one shown here is Escherichia coli , the true “workhorse” of molecular biologists. Its length is approximately 5 μm. Based on these dimensions and shapes of the cells shown, the bacterial cell is approximately 1/500th of the volume of the eukaryotic cell shown. Virus particles also vary greatly in size and shape, but generally range from 25 to 200 nm (0.25–2.00 × 10 −7m).
Infections with many viruses completely convert the cell into a factory for replication of new viruses. Under certain circumstances and/or in particular cells, however, virus infection leads to a state of coexistence between the cell and infecting virus, which can persist for as long as the life of the host. This process can be a dynamic one in which there is a small amount of virus produced constantly, or it can be passive where the viral genome is carried as a “passenger” in the cell with little or no evidence of viral gene expression. Often in such a case, the virus induces some type of change in the cell so that the viral and cellular genomes are replicated in synchrony. Such coexistence usually results in accompanying changes to the protein composition of the cell's surface – the immune “signature” of the cell – and often there are functional changes as well. This process is called lysogenyin bacterial cells and transformationin animal and plant cells.
In animal cells, the process of transformation often results in altered growth properties of the cell and can result in the generation of cells that have some or many properties of cancer cells. There are instances, however, where the coexistence of a cell and an infecting virus leads to few or no detectable changes in the cell. For example, herpes simplex virus ( HSV )can establish a latent infectionin terminally differentiated sensory neurons. In such cells there is absolutely no evidence for expression of any viral protein at all. Periods of viral latency are interspersed with periods of reactivation (recrudescence)where virus replication is reestablished from the latently infected tissue for varying periods of time.
Some viral infections of plant cells also result in stable association between virus and cell. Indeed, the variegation of tulip colors, which led to economic booms in Holland during the sixteenth century, is the result of such associations. Many other examples of mosaicismresulting from persisting virus infections of floral or leaf tissue have been observed in plants. However, many specific details of the association are not as well characterized in plants as in animal and bacterial cells.
Stages of virus replication in the cell
Various patterns of replication as applied to specific viruses, as well as the effect of viral infections on the host cell and organism, are the subject of many of the following chapters in this book. The best way to begin to understand patterns of virus replication is to consider a simple general case: the productive infectioncycle – this is shown schematically in Figure 2.2. A number of critical events are involved in this cycle. The basic pattern of replication is as follows:
1 The virus specifically interacts with the host cell surface, and the viral genome is introduced into the cell. This involves specific recognition between virus surface proteins and specific proteins on the cell surface (receptors) in animal and bacterial virus infections.
2 Viral genes are expressed using host cell processes. This viral gene expression results in synthesis of a few or many viral proteins involved in the replication process.
3 Viral proteins modify the host cell and allow the viral genome to replicate using host and viral enzymes. While this is a simple statement, the actual mechanisms by which viral enzymes and proteins can subvert a cell are manifold and complex. This is often the stage at which the cell is irreversibly modified and eventually killed. Much modern research in the molecular biology of virus replication is directed toward understanding these mechanisms.
4 New viral coat proteins assemble into capsids, and viral genomes are included. The process of assembly of new virions is relatively well understood for many viruses. The successful description of the process has resulted in a profound linkage of knowledge about the principles of macromolecular structures, the biochemistry of protein–protein and protein–nucleic acid interactions, and the thermodynamics of large macromolecule structures. Figure 2.2The virus replication cycle. Most generally, virus replication can be broken into the stages shown: (a) initial recognition between virus and cell and introduction of viral genetic material into the host cell, (b) virus gene expression and induction of virus‐induced modification of host, allowing (c) virus genome replication. Following this, (d) virus‐associated proteins are expressed, and (e) new virus is assembled and released, often resulting in cell death.
5 Virus is released where it can infect new cells and repeat the process. This is the basis of virus spread, whether from cell to cell or from individual to individual. Understanding the process of virus release requires knowledge of the biochemical interactions between cellular organelles and viral structures. Understanding the process of virus spread between members of a population requires knowledge of the principles of epidemiology and public health.
PATHOGENESIS OF VIRAL INFECTION
Most cells and organisms do not passively submit to virus infection. As noted in Chapter 1, the response of organisms to virus infection is a major feature of evolutionary change in its most general sense. As briefly noted, a complete understanding of pathogenesis requires knowledge of the sum total of genetic features a virus encodes that allows its efficient spread between individual hosts and within the general population of hosts. Thus, the term pathogenesis can be legitimately applied to virus infections of multicellular, unicellular, and bacterial hosts.
A major challenge for viruses infecting bacteria and other unicellular organisms is finding enough cells to replicate in without isolating themselves from other populations of similar cells. In other words, they must be able to “follow” the cells to places where the cells can flourish. If susceptible cells can isolate themselves from a pathogen, it is in their best interest to do so. Conversely, the virus, even when constrained to confine all its dynamic features of existence to the replication process per se, must successfully counter this challenge or it cannot survive.
Читать дальшеИнтервал:
Закладка:
Похожие книги на «Basic Virology»
Представляем Вашему вниманию похожие книги на «Basic Virology» списком для выбора. Мы отобрали схожую по названию и смыслу литературу в надежде предоставить читателям больше вариантов отыскать новые, интересные, ещё непрочитанные произведения.












Обсуждение, отзывы о книге «Basic Virology» и просто собственные мнения читателей. Оставьте ваши комментарии, напишите, что Вы думаете о произведении, его смысле или главных героях. Укажите что конкретно понравилось, а что нет, и почему Вы так считаете.