Pathology of Genetically Engineered and Other Mutant Mice
Здесь есть возможность читать онлайн «Pathology of Genetically Engineered and Other Mutant Mice» — ознакомительный отрывок электронной книги совершенно бесплатно, а после прочтения отрывка купить полную версию. В некоторых случаях можно слушать аудио, скачать через торрент в формате fb2 и присутствует краткое содержание. Жанр: unrecognised, на английском языке. Описание произведения, (предисловие) а так же отзывы посетителей доступны на портале библиотеки ЛибКат.
- Название:Pathology of Genetically Engineered and Other Mutant Mice
- Автор:
- Жанр:
- Год:неизвестен
- ISBN:нет данных
- Рейтинг книги:3 / 5. Голосов: 1
-
Избранное:Добавить в избранное
- Отзывы:
-
Ваша оценка:
Pathology of Genetically Engineered and Other Mutant Mice: краткое содержание, описание и аннотация
Предлагаем к чтению аннотацию, описание, краткое содержание или предисловие (зависит от того, что написал сам автор книги «Pathology of Genetically Engineered and Other Mutant Mice»). Если вы не нашли необходимую информацию о книге — напишите в комментариях, мы постараемся отыскать её.
An updated and comprehensive reference to pathology in every organ system in genetically modified mice Pathology of Genetically Engineered and Other Mutant Mice
Pathology of Genetically Engineered and Other Mutant Mice
Pathology of Genetically Engineered and Other Mutant Mice — читать онлайн ознакомительный отрывок
Ниже представлен текст книги, разбитый по страницам. Система сохранения места последней прочитанной страницы, позволяет с удобством читать онлайн бесплатно книгу «Pathology of Genetically Engineered and Other Mutant Mice», без необходимости каждый раз заново искать на чём Вы остановились. Поставьте закладку, и сможете в любой момент перейти на страницу, на которой закончили чтение.
Интервал:
Закладка:
Examination of the spleen: Routine examination is usually performed on two H&E‐stained cross sections of the spleen. Immunohistochemistry is performed to identify specific regions of the white pulp such as the marginal zone, and specific cell populations.
Lymphoid hypoplasia: Similar to the lymph nodes, mutations that affect the production of lymphocytes result in greatly decreased numbers of lymphocytes in the white pulp. Null mutations of Foxn1 cause a marked decrease of lymphocytes in the periarteriolar lymphocyte sheath ( Figure 7.6). Mutations of the Rag1, Rag2, and Prkdc genes result in a lack of B and T cells, and the white pulp is markedly reduced in size and largely devoid of lymphocytes ( Figure 7.6). Il2rg mutations affect the production of lymphocytes as well as other immune cells. NOD.Cg‐Prkdcscid Il2rgtm1Wjl/SzJ (NSG) mice have very small rudimentary white pulp ( Figure 7.6).
Aging‐associated changes: The white pulp of older mice (>12 months of age) becomes increasingly disorganized with less clear separation of B and T cell areas [55]. B cell follicles are reduced in size and there is a reduction in the number and function of follicular dendritic cells. The marginal zone and metallophilic marginal zone macrophages no longer form a continuous cell layer [55, 56].
Amyloidosis: Amyloid in the spleen is usually AA‐amyloid associated with chronic inflammation. Experimental models indicate that AA amyloid first accumulates in the marginal zone and is associated with loss of marginal zone macrophages [57]. As the amyloidosis increases in severity, deposits expand into the red pulp.
Pigment: The appearance of hemosiderin pigment is normal in the red pulp of the spleen. Red pulp macrophages are engaged in erythrophagocytosis and frequently contain hemosiderin. The amount of hemosiderin is greater in female mice, but varies by mouse strain. Hemosiderin can be distinguished from other pigments by Prussian blue staining ( Figure 7.7). Lipofuscin‐ceroid pigment may be observed in red pulp macrophages, but is less common than hemosiderin. In mice with black fur, the dorsal one‐third of the spleen is often dark red to black as a result of accumulation of melanin. Light microscopically, dendritic melanocytes and melanin‐laden macrophages are present in the red pulp, and occasionally in the white pulp ( Figure 7.7). Figure 7.6 Spleen. Spleen of C57BL/6J (a), B6.Cg‐Foxn1nu (b), B6.129S7‐Rag1tm1Mom (c), and NOD.Cg‐Prkdcscid Il2rgtm1Wjl/SzJ (NSG) mice (d).
Increased extramedullary hematopoiesis (EMH): While modest EMH is normal in the red pulp of the spleen of adult mice, increased EMH can lead to marked expansion of the red pulp and is a common cause of splenomegaly. The myeloid lineage predominates when there is inflammation in other organs, while hemorrhage and red blood cell destruction may lead to an increase of erythrocyte precursors and sometimes megakaryocytes.
Red pulp atrophy and hypoplasia: A reduced size of the red pulp may be seen in mice with blocks in cell division as a result of genetic mutations or chemical treatment.
Milky Spots
Milky spots are collections of leukocytes located just beneath the mesothelium in the omentum [58]. They are predominantly composed of macrophages and B1 B cells with fewer classical B2 B cells. They contain few T cells and lack a distinct T cell area that is present in other secondary lymphoid organs. Furthermore, follicular dendritic cells are absent from milky spots. Lymphoid structures similar to milky spots are found in pericardial, mediastinal, and mesenteric fat, and these are known as fat‐associated lymphoid clusters (FALCs) [59]. The formation of milky spots and FALCs requires LTA and CXCL13, but is independent of lymphoid tissue‐inducer cells. The number and size of milky spots and FALCs is increased by exposure to microbiota, suggesting that these tissues have features in common with both secondary and tertiary lymphoid tissues. Milky spots lack afferent lymphatics, and antigens enter via fenestrations in the overlying mesothelium. Inflammation and infections in the peritoneal cavity lead to a rapid increase of the number and size of milky spots as a result of an influx of peritoneal macrophages and recruitment of cells from the blood circulation. Immune responses can involve both B1 cells which differentiate into IgM and IgA‐secreting plasma cells and conventional B and T cells.
Tertiary Lymphoid Organs
Organized lymphoid tissues can form at sites of chronic inflammation associated with autoimmune diseases, infections, and neoplasms ( Figure 7.8). Immunohistochemistry can identify distinct T and B cell areas with high‐endothelial venules, follicles with follicular dendritic cells and sometimes germinal centers. The same cytokines and chemokines that orchestrate the formation of secondary lymphoid organs play a role, but tertiary lymphoid organs are formed after birth and not at preselected sites [33]. The role of tertiary lymphoid organs in inflammation and cancer is not well established. Tertiary lymphoid organs associated with autoimmune inflammation are thought to contribute to the autoimmune response and be detrimental. However, tertiary lymphoid organs formed in the adventitial layer of the aorta in ApoE‐deficient mice (B6.129P2‐ Apoe tm1Unc/J) with atherosclerosis seem to have a beneficial effect as the atherosclerosis was more severe in mice in which the formation of the serosal lymphoid tissue was blocked [60]. The presence of tertiary lymphoid organs in and near tumors is thought to increase the response to immunotherapy and to be a positive prognostic indicator [61].
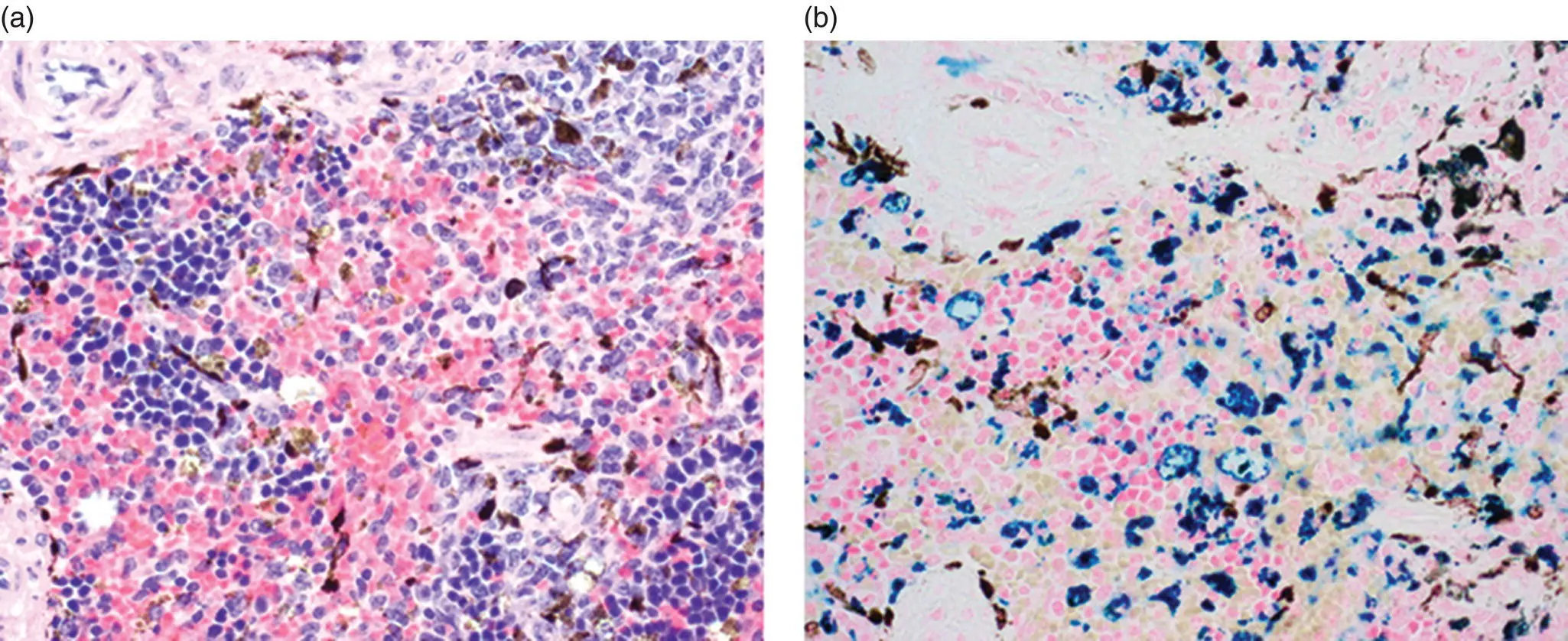
Figure 7.7 Melanin vs. hemosiderin. (a) Spleen of B6.Cg‐ Prkdc scid/SzJ mouse with melanocytes and melanin‐laden macrophages and hemosiderin. (b) Prussian blue stain distinguishes hemosiderin (blue) and melanin (dark brown) pigments.
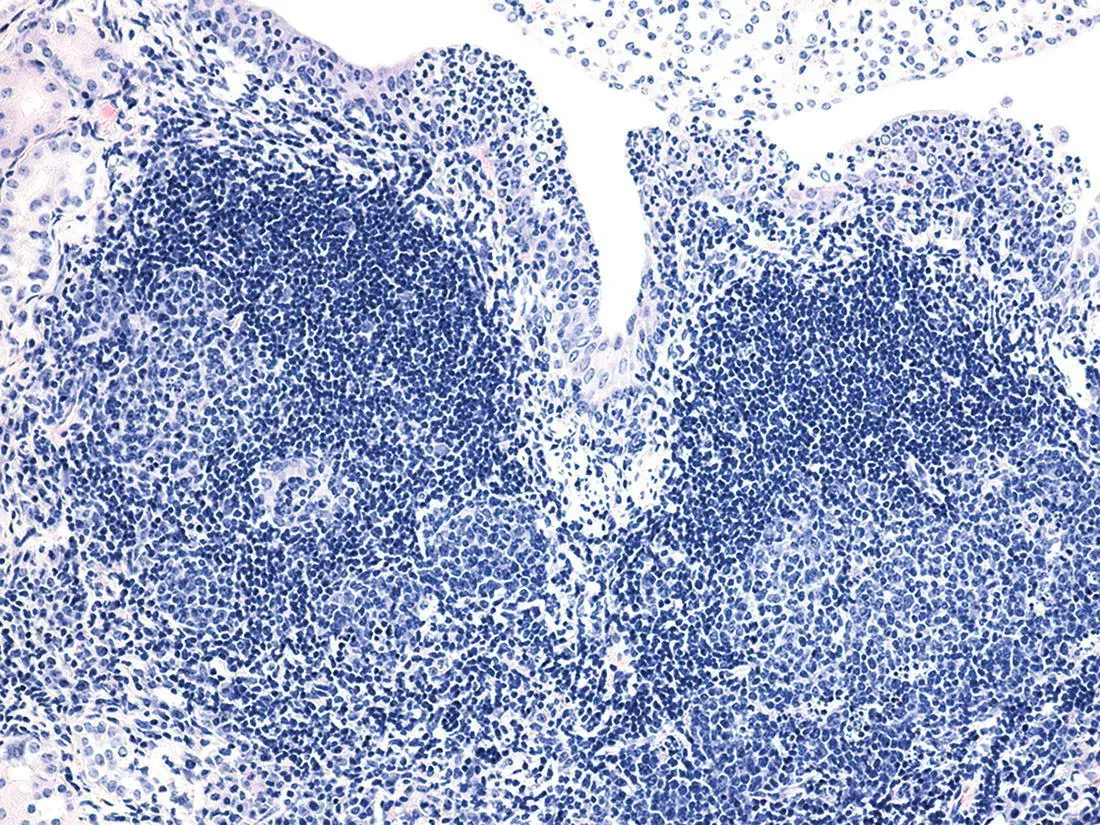
Figure 7.8 Tertiary lymphoid structures. Tertiary lymphoid tissue with two follicles with germinal centers in the kidney of a CD‐1 mouse with chronic pyelonephritis.
Acknowledgments
This work was supported grants from the National Institutes of Health (R01‐CA089713 and P30‐CA034196 to JPS).
References
1 1 Gao, X., Xu, C., Asada, N., and Frenette, P.S. (2018). The hematopoietic stem cell niche: from embryo to adult. Development 145 (2): dev139691.
2 2 Fitch, S.R., Kimber, G.M., Wilson, N.K. et al. (2012). Signaling from the sympathetic nervous system regulates hematopoietic stem cell emergence during embryogenesis. Cell Stem Cell 11 (4): 554–566.
3 3 Mariani, S.A., Li, Z., Rice, S. et al. (2019). Pro‐inflammatory aorta‐associated macrophages are involved in embryonic development of hematopoietic stem cells. Immunity 50 (6): 1439–1452. e5.
4 4 Willard‐Mack, C.L., Elmore, S.A., Hall, W.C. et al. (2019). Nonproliferative and proliferative lesions of the rat and mouse hematolymphoid system. Toxicol. Pathol. 47 (6): 665–783.
5 5 Pinho, S. and Frenette, P.S. (2019). Haematopoietic stem cell activity and interactions with the niche. Nat. Rev. Mol. Cell Biol. 20 (5): 303–320.
6 6 Pioli, P.D., Casero, D., Montecino‐Rodriguez, E. et al. (2019). Plasma cells are obligate effectors of enhanced myelopoiesis in aging bone marrow. Immunity 51 (2): 351–366. e6.
Читать дальшеИнтервал:
Закладка:
Похожие книги на «Pathology of Genetically Engineered and Other Mutant Mice»
Представляем Вашему вниманию похожие книги на «Pathology of Genetically Engineered and Other Mutant Mice» списком для выбора. Мы отобрали схожую по названию и смыслу литературу в надежде предоставить читателям больше вариантов отыскать новые, интересные, ещё непрочитанные произведения.












Обсуждение, отзывы о книге «Pathology of Genetically Engineered and Other Mutant Mice» и просто собственные мнения читателей. Оставьте ваши комментарии, напишите, что Вы думаете о произведении, его смысле или главных героях. Укажите что конкретно понравилось, а что нет, и почему Вы так считаете.